КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Другие вспомогательные белки поддерживают архитектуру миофибрилл и обеспечивают их эластичность
|
|
|
|
11-5
11-4
11-3
11.1.3. Сокращение - результат скольжения тонких и толстых филаментов друг относительно друга [2]
Если через живую мышечную клетку пропустить пучок монохроматического света, возникнет интерференционная картина, позволяющая с большой точностью регистрировать изменения в длине саркомеров. Такие измерения показали, что при сокращении мышцы пропорционально укорачивается и каждый саркомер; если миофибрилла, состоящая из 20000 саркомеров, укорачивается с 5 см до 4 см (т.е. на 20%), длина каждого саркомера соответственно уменьшится с 2,5 до 2 мкм.
При укорочении саркомера сжимаются только светлые полосы - темная полоса своих размеров не меняет. Это можно легко объяснить, предположив, что сокращение вызывается скольжением тонких филаментов относительно толстых без изменения длины тех и других (рис. 11-4).
Эта «.модель скользящих нитей», впервые предложенная в 1954 г., сыграла решающую роль в понимании механизма мышечного сокращения. Она, в частности, привлекла внимание к молекулярным взаимодействиям, лежащим в основе взаимного скольжения соприкасающихся толстых и тонких филаментов.
Модель скользящих нитей базируется на нескольких группах экспериментальных данных. Электронномикроскопические исследования показали, что длина как толстых, так и тонких филаментов при укорочении мышцы не изменяется. Судя по данным рентгеноструктурного анализа, характер упаковки субъединиц, образующих филаменты, тоже остается неизменным. По мере укорочения мышцы развиваемое механическое усилие растет пропорционально степени перекрывания толстых и тонких филаментов; этого и следует ожидать, если усиление - результат взаимодействия нитей во всей области их соприкосновения.
Ультраструктурную основу этого взаимодействия удается выявить с помощью электронной микроскопии высокого разрешения.
Оказалось, что от толстых филаментов отходят многочисленные боковые отростки, или поперечные мостики, соприкасающиеся с тонкими нитями, которые лежат на расстоянии около 13 нм от толстых (рис. 11-5). При сокращении мышцы толстые и тонкие филаменты подтягивают друг друга с помощью этих мостиков, работающих циклично, как миниатюрные весла.
Взаимодействующие белки тонких и толстых филаментов были идентифицированы как актин и миозин соответственно. Актин, которого в цитоскелете больше, чем какого-либо другого белка, часто образует вместе с миозином структуры, способные к сокращению. Хотя эти белки
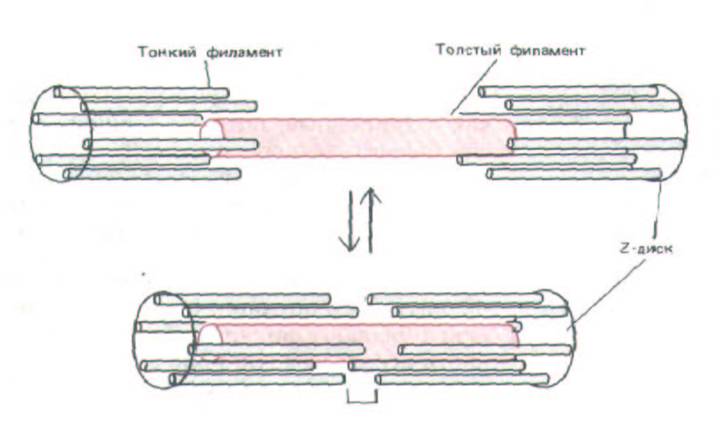
Рис. 11-4. Схема, иллюстрирующая процесе мышечного сокращения по принципу скользящих нитей; толстые и тонкие филаменты скользят друг по другу, не изменяя собственной длины.
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.

Рис. 11-5. Продольный срез летательной мышцы насекомого (электронная микрофотография; препарат получен методом быстрого замораживания, скалывания и глубокого травления). Обратите внимание на почти кристаллическую укладку толстых миозиновых и тонких актиновых филаментов. Поперечные мостики, соединяющие нити двух типов, - головки миозина. (С любезного разрешения John Heuser и Roger Cooke.)
имеются почти во всех эукариотических клетках, большая часть наших знаний об их свойствах первоначально была получена в биохимических экспериментах с актином и миозином, выделенными из мышцы.
11.1.4, Тонкие филаменты состоят в основном из актина [2]
Актин имеется у всех эукариот, включая одноклеточных (например, у дрожжей). Гены актина эволюционно крайне консервативны, так что актины весьма далеких друг от друга организмов в опытах in vitro функционально взаимозаменяемы. Главные свойства актина, выделенного, например, из скелетных мышц позвоночных, являются общими| для актинов из любых других источников.
Обычно актин выделяют, обрабатывая порошок высушенной мышечной ткани сильно разбавленным солевым раствором, который вызывает диссоциацию актиновых филаментов на их глобулярные субъединицы. Каждая субъединица представляет собой одну полипептидную цепь длиной в 375 аминокислотных остатков, с которой нековалентно связана одна молекула АТР. Такой актин называют глобулярным, или G-
актином. При полимеризации актина связанный АТР гидролизуется, отщепляя концевой фосфат, а актин образует филаменты, называемые фибриллярным актином (F-актином). Полимеризацию можно вызвать, просто повысив концентрацию соли до уровня, близкого к физиологическому; при этом раствор актина, лишь ненамного более вязкий, чем вода, быстро «густеет» по мере образования филаментов.
Хотя в процессе полимеризации и происходит гидролиз связанного АТР, сама полимеризация энергии не требует; она идет, даже если. с актином связан ADP или негидролизуемый аналог АТР. Однако гидролиз АТР оказывает существенное влияние на динамическое поведение актиновых филаментов; это мы увидим позже, когда будем рассматривать те виды клеточной активности, которые (в отличие от мышечного сокращения) зависят от контролируемой полимеризации и деполимеризации актина.
На электронных микрофотографиях актиновые филаменты выглядят как однородные нити толщиной около 8 нм (рис. 11-6). Эти нити составляют основу тонких филаментов скелетных мышц, что подтверждается данными электронной микроскопии, рентгеноструктурного анализа и окраски антителами к актину. Однако актин - не единственный компонент тонких филаментов, о чем будет сказано позже (разд. 11.1.12).
Актиновые филаменты представляют собой плотную спираль, собранную из однотипно ориентированных мономеров актина (рис. 11-7).
Они обладают полярностью, т. е. два их конца различны. Эта полярность играет важную роль в осуществлении подвижности клеток и легче всего обнаруживается в ориентированных комплексах, которые каждый
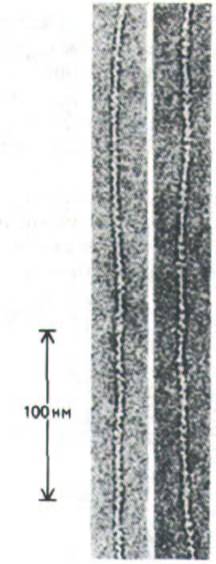
Рис. 11-6. Электронные микрофотографии актиновых филаментов (негативное контрастирование). {С любезного разрешения Roger Craig.)
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
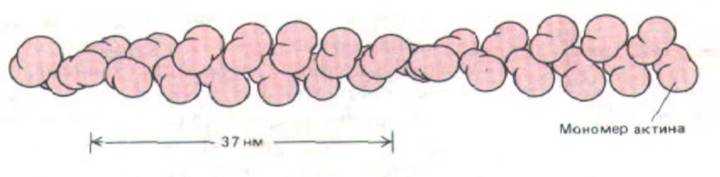
Рис. 11-7. Организация глобулярных молекул актина в актиновом филаменте. Молекулы упакованы в плотную спираль; на один оборот приходится приблизительно два мономера актина. Хотя такое устройство создает видимость спирали из двух цепочек актина, обвивающих друг друга с шагом 37 нм, эта видимость ошибочна, так гипотетическая "одиночная" актиновая цепь сама по себе существовать не может.
актиновый мономер образует с миозином. Но прежде чем обсуждать это ключевое взаимодействие, мы должны рассмотреть некоторые особенности молекул миозина.
11.1.5. Толстые филаменты состоят из миозина [2]
Миозин есть почти во всех клетках позвоночных и всегда находится в сократительных пучках, образуемых в цитоплазме актиновыми филаментами. Миозин - эволюционно гораздо менее консервативный белок, чем актин, и известно несколько его форм. При полимеризации in vitro миозин скелетных мышц, например, образует значительно более крупные филаменты, чем миозины немышечных клеток.
Миозин экстрагируют из скелетных мышц концентрированными солевыми растворами, под действием которых толстые филаменты деполимеризуются до составляющих их молекул миозина (рис. 11-8). Каждая молекула состоит из шести полипептидных цепей - двух одинаковых тяжелых цепей и двух пар легких цепей (рис. 11-9).
Протеолитический фермент папаин расщепляет молекулу миозина на длинный α-спиральный участок, называемый миозиновым
стержнем (или миозиновым хвостом), и две раздельные глобулярные миозиновые головки, называемые также субфрагментами-1 или S1-
фрагментами (рис. 11-10). Эти две части молекулы выполняют разные функции - хвост ответствен за самопроизвольную сборку толстых филаментов, а с помощью головок осуществляется движение этих филаментов относительно прилегающих актиновых нитей. Вначале мы опишем строение и самосборку хвостов, а затем рассмотрим, каким образом головки создают мышечное усилие.
11.1.6. Миозиновые хвосты самоорганизуются в биполярные толстые филаменты [3]
Подобно многим другим цитоскелетным белкам, хвосты миозина - это длинные стержневидные образования. Жесткость таких белков определяется наличием общего структурного элемента, в котором две α-спирали благодаря особому расположению гидрофобных аминокислотных остатков обвиваются друг около друга, образуя «скрученную спираль» (рис. 11-11). В миозине и во многих других белках цитоскелета эти две спирали направлены параллельно (ориентация N- и С-концов у них совпадает) и объединены в нить толщиной около 2 нм.
В то время как структура отдельных молекул миозина определяется
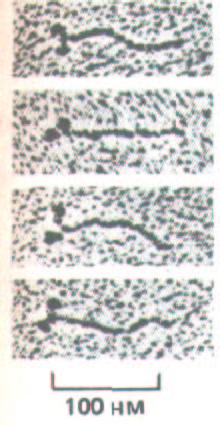
Рнс. 11-8. Электронные микрофотографии молекул миозина (напыление платиной). Обратите внимание, что каждая молекула состоит из двух глобулярных головок, прикрепленных к фибриллярному хвосту (С любезного разрешения David Shotton.)
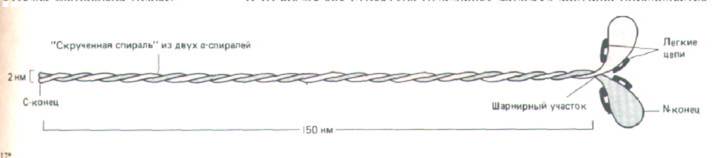
Рнс. 11-9. Молекула миозина построена из двух тяжелых цепей (каждая длиной около 2000 аминокислотных остатков) и четырех легких цепей. Легкие цепи представлены молекулами двух типов (в одних около 190 аминокислотных остатков, в других около 170)-по одной молекуле каждого типа в каждой миозиновой головке.
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
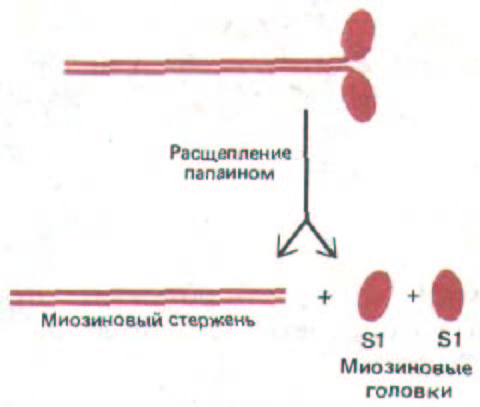
Рис. 11-10. При ограниченном расщеплении папаином молекула миозина распадается на стержень и две головки.
гидрофобными взаимодействиями между двумя α-спиральными тяжелыми цепями (рис. 11 -11, Л), структура толстых филаментов, образуемых миозином в мышце, зависит от ионных взаимодействий между хвостами. Именно поэтому растворы соли высокой концентрации, разрушающие ионные взаимодействия, но не влияющие на гидрофобные, экстрагируют из мышцы отдельные молекулы миозина. При снижении ионной силы раствора до физиологического уровня эти молекулы ассоциируют, образуя крупные филаменты, которые могут быть очень сходны с нормальными толстыми филаментами мышц. В мышечных клетках эти взаимодействия стабилизируются различными сопутствующими белками, и получающиеся в результате толстые филаменты образованы сотнями миозиновых хвостов, упакованных в плотные упорядоченные пучки, из которых торчат миозиновые головки расположенные «лесенкой» (рис. 11-12). Такая структура оказывается биполярной, с «голой» (без миозиновых головок) центральной областью, где соединяются противоположно направленные пучки миозиновых хвостов. Глобулярные головки миозина взаимодействуют с актином, образуя поперечные мостики между толстыми и тонкими филаментами.
11.1.7. Источником энергии для мышечного сокращения служит гидролиз АТР [4]
Скелетная мышца превращает химическую энергию в механическую работу с весьма высокой эффективностью - в виде тепла теряется всего лишь 30-50% (для сравнения: тепловые потери при работе автомобильного двигателя составляют обычно 80-90% всей энергии, выделяющейся при сжигании бензина).
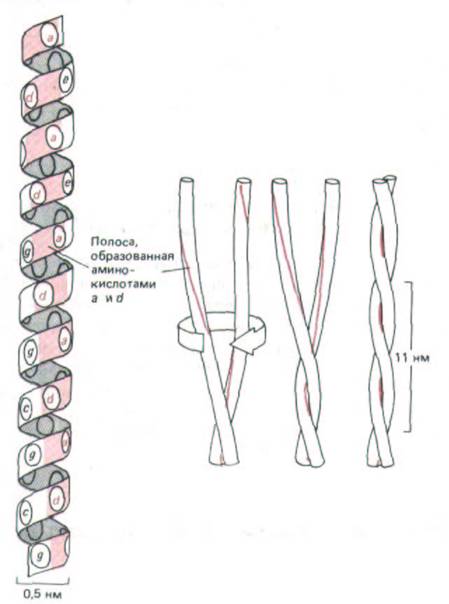
Рис. 11-11. Топология «скрученной спирали». Слева одиночная α-спираль представлена в виде цилиндра, где боковые цепи аминокислот обозначены семичленной последовательностью букв abcdefg (снизу вверх). Аминокислоты а и d в этой последовательности оказываются на поверхности цилиндра рядом, образуя «полосу» (выделена цветом), которая медленно оборачивается вокруг α -спирали. Белки, образующие скрученную спираль, как правило, имеют в положениях а и d гидрофобные аминокислоты. Поэтому, как показано справа, две α -спирали обвивают друг друга таким образом, что гидрофобные боковые цепи одной α -спирали попадают в пространство между гидрофобными боковыми цепями другой, тогда как более гидрофильные боковые цепи обращены к окружающей водной среде.
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.

Рис. 11-12. Толстый миозиновый филамент. А. Электронная микрофотография толстого филамента из мышцы морского гребешка. Видна центральная «голая» зона. Б. Схема строения (без соблюдения масштаба). Молекулы миозина связаны хвостовыми участками в пучок, на поверхности которого выступают головки. «Голая» зона в центре содержит только хвосты миозина. В. Небольшой отрезок толстого филамента:
реконструкция по электронным микрофотографиям. Одна из молекул миозина выделена цветом. (А-с любезного разрешения R. Craig; В-no R. A.
Crowther, R. Pardon, R. Craig, J. Моl, Biol. 184: 429-439, 1985.)
Необходимую для мышечного сокращения энергию поставляет гидролиз АТР, однако содержание АТР в покоящейся и в активно работающей мышце различается мало, так как в мышечных клетках работает чрезвычайно эффективная система регенерации АТР. Фермент фосфокреатинкиназа катализирует реакцию между креатинфосфатом и ADP, в результате которой образуются АТР и креатин. (Креатин-фосфат-
вещество с еще большей энергией, чем АТР; рис. 11-13). После кратковременной вспышки мышечной активности падает внутриклеточный уровень именно креатинфосфата, хотя сам сократительный механизм использует АТР. Таким образом, креатинфосфат играет роль аккумулятора - он запасает энергию, «заряжаясь» за счет новых молекул АТР (синтезируемых при клеточных процессах окисления), когда мышца находится в покое.
11.1.8. Миозин действует как актин-зависимая АТРаза [5]
Происходящий при мышечном сокращении гидролиз АТР - прямое следствие взаимодействия между актином и миозином. Миозин и сам по себе действует как АТРаза, но в очищенном виде он работает сравнительно медленно. Для завершения полного цикла гидролиза одной молекулы АТР каждой молекуле миозина требуется примерно полминуты. При этом скорость-лимитирующей стадией оказывается не связывание АТР с миозином и не гидролиз концевой фосфатной связи (оба процесса протекают быстро), а освобождение продуктов гидролиза-ADP и неорганического фосфата - из комплекса с миозином. Оставаясь нековалентно связанными с его молекулой, они препятствуют присоединению и последующему гидролизу новых молекул АТР.
В присутствии актиновых филаментов АТРазная активность миозина резко возрастает. Каждая молекула миозина начинает гидролизовать от 5 до 10 молекул АТР в секунду, что сравнимо со скоростью гидролиза в сокращающейся мышце. Активация АТРазы миозина актиновыми филаментами отражает физическую ассоциацию этих белков, которая служит основой мышечного сокращения. Связывание миозина с актиновым филаментом ведет к быстрому отделению ADP и PI от молекулы миозина, в результате чего последняя может связывать новые молекулы АТР и возобновлять цикл.
11.1.9. С актиновыми филаментами взаимодействуют головки миозина [6]
Как связывание с актиновыми филаментами, так и гидролиз АТР осуществляют глобулярные миозиновые головки. Изолированные го-
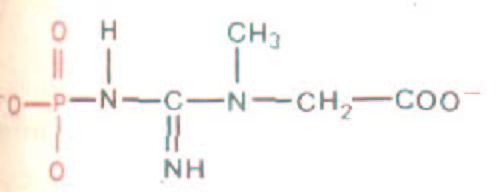
Рис. 11-13. Структура креатинфосфата. Это вещество играет роль «запасного аккумулятора» высоко-.нергетических фосфатных групп в мышцах и других тканях позвоночных. Фосфатная группа (выделена цветом) с помощью фермента греатинкиназы переносится на ADP с образованием АТР, когда в последнем возникает нужда.
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
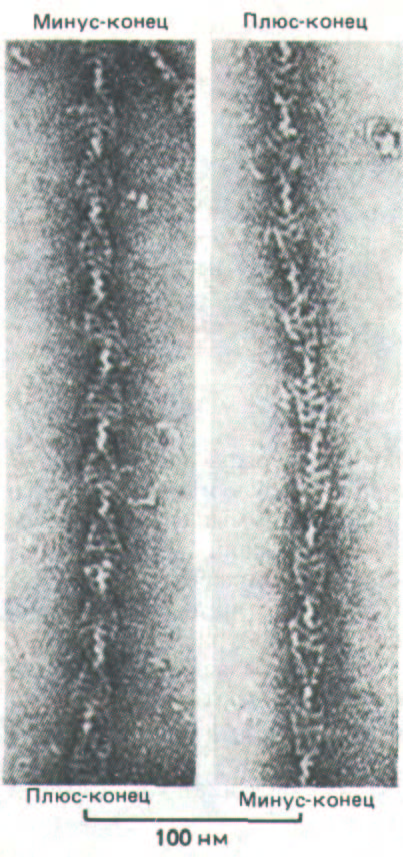
Рис. 11-14. Электронная микрофотография актиновых филаментов с присоединенными к ним изолированными головками миозина.
Спиральное расположение связанных головок, повернутых в одном направлении, создает картину цепочки из наконечников стрел, выявляющую полярность актинового филамента. Конец, к которому обращены острия, называют минус-концом, а другой конец - плюс-концом, так как полимеризация актина на этих концах идет с различной скоростью (см. рис. 11-40). {С любезного разрешения R. Craig.)
ловки, которые можно получить путем отщепления «хвостов» папаином (см. рис. 11-10), сохраняют и АТРазную активность, и актин-связывающие свойства интактных молекул миозина, поэтому их можно использовать для изучения взаимодействий между актином и миозином.
Каждая молекула актина в составе актинового филамента способна связать одну миозиновую головку. Образующиеся при этом комплексы выдают структурную полярность актиновых филаментов; в электронном микроскопе негативно контрастированные препараты таких филаментов имеют весьма характерный вид: каждая миозиновая головка образует боковой выступ, и все множество этих выступов создает впечатление, что на филамент нанизаны наконечники стрел (рис. 11-14) Поскольку миозиновые головки присоединяются к каждой субъединице актина в одинаковой ориентации, такая картина означает, что все актиновые молекулы тоже ориентированы вдоль оси филамента в одном направлении. Таким образом, два конца актинового филамента структурно различаются. Их назвали соответственно минус-концом (или заостренным концом, т. е. тем, к которому направлены острия стрел) и плюс-концом (или оперенным концом, к которому обращены хвосты стрел).
Термины «плюс» и «минус» связаны с тем фактом, что разные концы актинового филамента in vitro растут с различной скоростью (разд. 11.20.9).
Из рисунка 11-12 видно, что миозиновые головки, расположенные по разные стороны от центральной «голой» области толстого филамента, смотрят в противоположных направлениях. Так как головки должны взаимодействовать с тонкими филаментами в зоне перекрывания, сами тонкие филаменты с одной и с другой стороны саркомера должны иметь противоположную полярность. Это действительно удалось продемонстрировать, присоединяя миозиновые головки к актиновым филаментам, отходящим в обе стороны от изолированных Z-дисков: все миозиновые «стрелы» были направлены прочь от Z-диска. Таким образом, плюс-концы каждого актинового филамента закреплены в Z-диске, а минус-концы направлены в сторону толстых филаментов (рис. 11-15).
11.1.10. Миозиновые головки «шагают» по актиновому филаменту в направлении плюс-конца [7]
Мышца сокращается в результате взаимодействия головок миозина с прилегающими к ним актиновыми филаментами. В ходе этого взаимодействия миозиновые головки гидролизуют АТР. Гидролиз АТР и последующая диссоциация прочно связанных продуктов гидролиза (ADP и РI) вызывают упорядоченную серию аллостерических изменений в конформации миозина. В итоге часть освобождающейся энергии превращается в двигательную работу. Общие принципы, лежащие в основе сопряжения гидролиза АТР с направленным перемещением белковых молекул, обсуждаются в разд. 3.4.11.
Анализ кинетики гидролиза АТР в процессе мышечного сокращения, данные электронной микроскопии и результаты рентгеноструктурного анализа указывают на вероятную последовательность событий, представленную на рис. 11-16. Свободная головка миозина связывает АТР (состояние 1) и гидролизует его. Этот процесс обратим, так как энергия гидролиза АТР первоначально запасается в напряженной конформации белка (когда ADP и PJ остаются связанными с ним - состояние 2). Переходя поочередно в то или другое из этих состояний, миозиновая головка в результате случайных движений может приблизиться к соседней субъединице актина и слабо связаться с ней; это приводит к освобождению PI, что в свою очередь ведет к прочному связыванию головки с актиновым филаментом (состояние 3). В этом состоянии головка
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
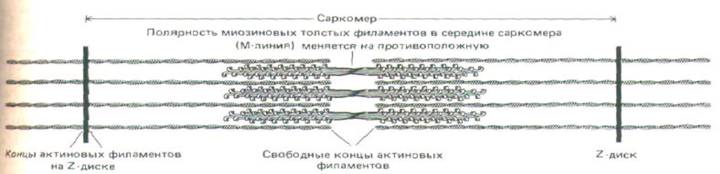
Рис. 11-15. Тонкие и толстые филаменты в саркомере перекрываются, причем их полярность симметрична относительно средней линии саркомера.
претерпевает конформационное изменение, которое и производит элементарный силовой акт, участвующий в подтягивании всего толстого филамента. В конце этого «рабочего хода» (состояние 4) происходит отделение ADP от головки, а затем присоединение к ней новой молекулы АТР, отделяющее головку от актинового филамента и возвращающее ее в состояние 1. Далее гидролиз связанного АТР подготавливает миозиновую головку к следующему циклу.
Поскольку каждый оборот цикла приводит к гидролизу и освобождению одной молекулы АТР, вся эта серия конформационных изменений связана с большим положительным изменением свободной энергии системы, что делает весь процесс однонаправленным (см. разд.
3.4.11). Поэтому каждая отдельная головка миозина «шагает» по актиновому филаменту в одном направлении - всегда в сторону плюс-концов (см.
рис. 11-15). При циклических изменениях своей конформации головка тянет актиновый филамент, заставляя его скользить относительно миозинового; в те моменты, когда головка не контактирует с актином, она
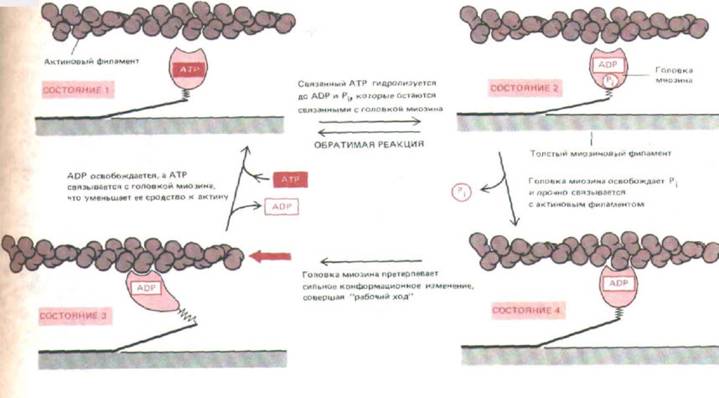
Рис. 11-16. Эта схема показывает, какнм образом молекула миозина может использовать энергию гидролиза АТР, чтобы двигаться по актвому филаменту от его минус-конца к плюс-концу. При переходе из состояния 2 в состояние 3 присоединение миозиновой голов-в к актину приводит к тому, что она теряет связанный с ней фосфат и более прочно прикрепляется актиновому филаменту. Вслед за этим форма головки претерпевает пока еще не очень понятные изменения, которые сопровождаются высвобождением ADP и заставляют миозиновую головку «подтянуться» относительно актинового филамента (рабочий ход). Каждая из пары головок на молекуле миозина работает независимо от другой.
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
смещается за счет работы других миозиновых головок того же филамента. На «моментальном снимке» всего миозинового филамента в сокращающейся мышце было бы видно, что часть его головок находится в контакте с актиновыми филаментами, а часть остается свободной (для этого весьма существенна эластичность, «пружинистость» молекулы миозина). На каждом толстом филаменте сидит около 500 миозиновых головок, и каждая головка при быстром сокращении мышцы проходит примерно 5 рабочих циклов за секунду; в результат скорость скольжения тонких филаментов относительно толстых достигает 15 мкм/с.
11.1.11. Мышечное сокращение инициируется внезапным повышением концентрации Са2+ в цитозоле [8]
Только что описанный молекулярный механизм создания силы включается лишь тогда, когда мышца получает сигнал от своего мотонейрона. Нервный импульс вызывает на плазматической мембране мышечной клетки потенциал действия, и в результате электрическое возбуждение быстро распространяется по серии мембранных впячиваний, называемых поперечными трубочками (Т-трубочками), которые отходят внутрь от плазматической мембраны, вступая в контакт с каждой миофибриллой. Отсюда сигнал каким-то образом передается саркоплазматическому ретикулуму - своеобразной оболочке из сообщающихся уплощенных пузырьков, которая окружает каждую миофибриллу подобно сетчатому чулку (рис. 11-17).
Щель между Т-трубочкой и саркоплазматическим ретикулумом имеет ширину всего лишь 10-20 нм, но как через нее передается сигнал, остается неясным. Электрическое возбуждение Т-трубочек приводит (неизвестным пока способом) к открытию в мембране саркоплазматического ретикулума кальциевых каналов (рис. 11-17), и ионы Са2+, которых в ретикулуме очень много, выходят в цитозоль. Именно этот внезапный подъем концентрации свободных ионов Са2+ в цитозоле вызывает сокращение миофибрилл. Так как время прохождения сигнала от плазматической мембраны через Т-трубочки и саркоплазматический ретикулум до каждого из саркомеров измеряется миллисекундами, сокращение всех миофибрилл в клетке происходит одновременно. Повышение концентрации Са2+ в цитозоле кратковременно, поскольку ионы Са2+ быстро перекачиваются обратно в саркоплазматичес-
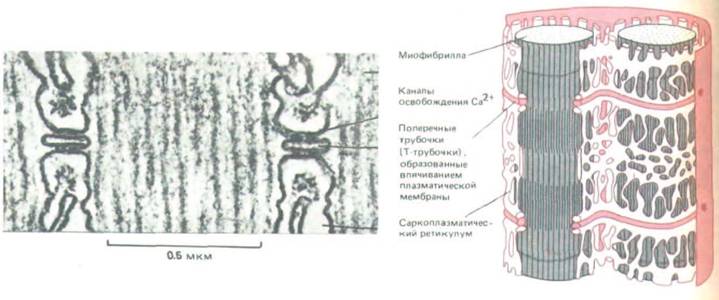
Рис. 11-17. Система мембран, передающая сигнал к сокращению от плазматической мембраны мышечной клетки ко всем ее миофибриллам. На электронной микрофотографии видны две Т-трубочки и большие каналы для выброса Са2+ в мембране саркоплазматического ретикулума, которые выглядят как прямоугольные «ножки», соединенные с мембраной соседней Т-трубочки. (Микрофотографию любезно предоставила Clara Ftanzini-Armstrong.)
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
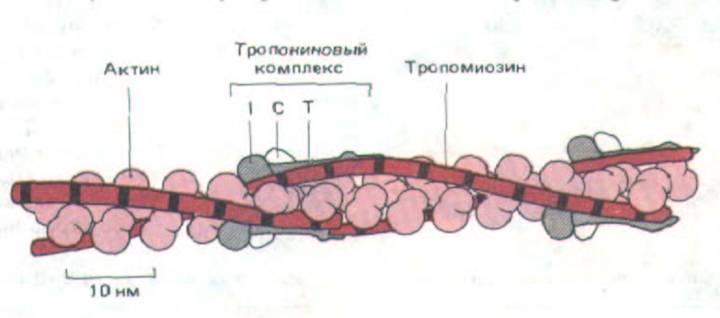
Рис. 11-18. Здесь показано, каким образом на актиноном филаменте расположены тропомиозин и тропонин. Каждая молекула тропоми-
озина имеет семь регулярно расположенных участков с гомологичной последовательностью; как полагают, каждый такой участок связывается с мономером актина. Обратите внимание, что концы соседних молекул тропомиозина слегка перекрываются, как если бы они полимеризовались вдоль актинового филамента «головой к хвосту». (По G.N. Phillips, J.P. Fillers, С. Cohen, J. Моl. Biol. 192: 111-131, 1986, с изменениями.)
кий ретикулум Са2 + -АТРазой, которой в его мембране очень много (разд. 6.4.7). Обычно возвращение концентрации Са2 + в цитозоле к уровню покоя происходит за 30 мс, что приводит к расслаблению миофибрилл.
11.1.12. Сокращение скелетной мышцы регулируется ионами Са2 + при участии тропонина и тропомиозина [9]
Зависимость сокращения скелетной мышцы позвоночного от ионов Са2 + (и тем самым - от команд, передаваемых нервами) всецело определяется наличием специализированных вспомогательных белков, тесно связанных с актиновыми филаментами. Если миозин смешать в пробирке с очищенными актиновыми филаментами, АТРаза миозина активируется независимо от присутствия Са2 +, тогда как в нормальной миофибрилле, где на актиновых филаментах «сидят» вспомогательные белки, активация миозиновой АТРазы зависит от Са2 +.
Один из этих белков, названный тропомиозином (из-за сходства его с миозином на рентгеновских дифрактограммах), - жесткая стержневидная молекула длиной 41 нм. Как и хвост миозина, тропомиозин представляет собой димер, состоящий из двух одинаковых α-
спиральных цепей по 284 аминокислоты в каждой. Цепи обвиваются друг около друга (см. разд. 11.1.6). Связываясь с актиновым филаментом по всей его длине, он стабилизирует филамент и придает ему жесткость (рис. 11-18).
Другой важный вспомогательный белок, участвующий в регулировании функции скелетных мышц позвоночных ионами кальция, - это тропонин. Он представляет собой комплекс из трех полипептидов-тропонинов Т, I и С (названных так за свои тропомиозин-связывающие, ингибиторные и Са2+-связывающие свойства соответственно). Тропониновый комплекс имеет удлиненную форму, причем субъединицы С и I образуют глобулярную головку, а тропонин Т-длинный хвост. Хвост тропонина Т связывается с тропомиозином и, как полагают, определяет положение всего комплекса на тонком филаменте (рис. 11-18). Тропонин I присоединяется к актину, и если его добавить к тропонину Т и тропомиозину, то образуемый ими комплекс будет ингибировать взаимодействие актина с миозином даже в присутствии Са2+.
Если же, наконец, добавить тропонин С и достроить таким образом тропониновый комплекс, его влияние на актин-миозиновые взаимодействия станет чувствительным к Са2+. Тропонин С способен связывать до четырех ионов Са2+, и в комплексе с Са2+ он ослабляет ингибирующее действие двух других компонентов тропонина на связывание миозина с актином. Тропонин С-близкий «родственник» кальмодулина
- белка, который участвует в Са2 +-зависимых ответах всех клеток, в том числе в активации миозина в гладких мышцах (разд. 11.1.15). Поэтому тропонин С можно рассматривать как специализи-
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
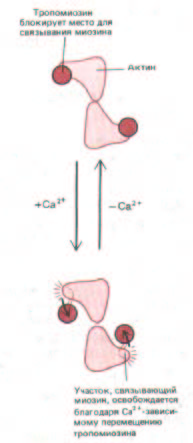
Рис. 11-19. Актиновый филамент в поперечном разрезе (схема): показано, каким образом в отсутствие Са2+ тропомиозин может блокировать взаимодействие миозиновых головок с актином.
рованную форму кальмодулина, приобретшую участки стабильного связывания с тропонином I и тропонином Т и обеспечивающую тем самым способность миофибрилл чрезвычайно быстро реагировать на повышение концентрации Са2 +.
На каждые семь мономеров актина в актиновом филаменте приходится только один тропониновый комплекс (рис. 11-18). Судя по данным структурных исследований, в покоящейся мышце связывание тропонина I с актином ведет к перемещению тропомиозина на актиновом филаменте в то самое место, с которым в сокращающейся мышце контактируют миозиновые головки, и в результате взаимодействие актина с миозином подавляется. При повышении уровня Са2 + тропонин С заставляет тропонин I «отцепиться» от актина, таким образом позволяя тропомиозину слегка изменить свое положение, и участок взаимодействия головок актина с миозином освобождается (рис. 11-19}.
[10]
Удивительная сила и быстрота мышечного сокращения обусловлена тем, что в каждой миофибрилле актиновые и миозиновые филаменты находятся на оптимальном расстоянии друг от друга и в правильном расположении. Тонкую организацию миофибрилл обеспечивает группа
| Таблица 11-1. Основные белковые компоненты миофибрилл скелетных мышц позвоночных1' | ||||
| Белок | Доля общего белка, % | Мол. масса | Субъединицы, мол. масса | . Функция |
| Миозин | 2 x 223 {тяжелые цепи) | Главный компонент толстых филаментов. Взаимодействует с актиновыми филаментами, создавая механическое усилие за счет гидролиза АТР | ||
| Актин | - | Главный компонент тонких филаментов, по которым скользят толстые филаменты при мышечном сокращении | ||
| Тропомиозин | 2x32 | Стержневидный белок, который связывается с актиновыми филаментами по всей их длине | ||
| Тропонин | 30 (Τn = Т) | Комплекс из трех мышечных белков, расположен через равные | ||
| 30(Тn = 1) | интервалы на актиновых филаментах и участвует в регуляции | |||
| 18 (Τn = С) | мышечного сокращения ионами Ca2 + | |||
| Титин | ~2500 | - | Очень большой гибкий белок, образует эластичную цепь, соединяющую толстые филаменты с Ζ- дисками | |
| Небулин | - | Удлиненный нерастяжимый белок, связанный с Z-диском и ориентированный параллельно актиновым филаментам | ||
| a-Актинин | 2x95 | Актин-связывающий белок, который соединяет актиновые филаменты в области Ζ- диска. | ||
| Миомезин | - | Миозин-связывающий белок, находящийся в области центральной М- линии толстых филаментов | ||
| С-белок | - | Миозин-связывающий белок; образует полосы по обе стороны от М- линии толстых филаментов. | ||
| 1) Поперечно полосатые миофибриллы позвоночных содержат также не менее 20 других белков, не вошедших в эту таблицу. |
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
Рис. 11-20. Электронная микрофотография молекул очищенного α-актинина (любезно предоставлена J. Heuser.)
структурных белков, из которых больше дюжины уже идентифицировано (табл. 11-1), Локализация большинства этих белков в саркомере была выяснена иммуноцитохимическими методами (разд. 4.5.3).
Актиновые филаменты «заякорены» своими плюс-концами в Z-диске, где их удерживают в правильно организованной решетке другие белки. Из них лучше всего охарактеризован а-актинин - актин-связывающий белок, имеющийся в большинстве животных клеток. В мышечных клетках он находится в области Z-диска. Очищенный а-актинин - биполярная палочковидная молекула (рис. 11-20), которая может связывать актиновые филаменты в параллельные пучки. Аналогичную функцию в случае миозина может выполнять белок миомезин, который сшивает соседние миозиновые филаменты в области М-линии (посередине биполярного толстого филамента), собирая их в гексагональную упаковку.
Стабилизирует упаковку миозиновых филаментов еще одна группа миозин-связывающих белков, выявляемых при окраске антителами как серия из 11 регулярно расположенных бледных полосок по обе стороны от М-линии.
В мышечных клетках есть также целая система очень плохо растворимых белковых филаментов, которые можно выделить лишь после полной экстракции миозина и актина из саркомера концентрированным раствором йодистого калия. Одна группа таких филаментов, построенных из очень крупного белка, названного титином, тянется параллельно толстым и тонким филаментам и соединяет толстые филаменты с Z-диском.
Титиновые филаменты очень эластичны и, по-видимому, действуют как пружины, «центрируя» толстые филаменты между Z-дисками (рис. 11-21).
Еще одна группа нерастворимых нитей - это промежуточные филаменты (разд. 11.5), которые расположены между Z-дисками соседних миофибрилл. Предполагается, что они удерживают саркомеры в определенных пространственных отношениях между собой и соединяют миофибриллы с плазматической мембраной мышечной клетки.
11.1.14. У позвоночных есть три основных типа мышц [11]
До сих пор мы рассматривали лишь один из трех главных типов мышц, имеющихся у позвоночных, а именно скелетные мышцы. Два других-это сердечная мышца, которая за среднее время жизни человека успевает совершить около трех миллиардов циклов сокращения и расслабления, и гладкая мускулатура, обеспечивающая более медленное и продолжительное сокращение, характерное для таких органов, как кишечник. Во всех трех типах мышечных клеток, а также в еще одном типе сократи-
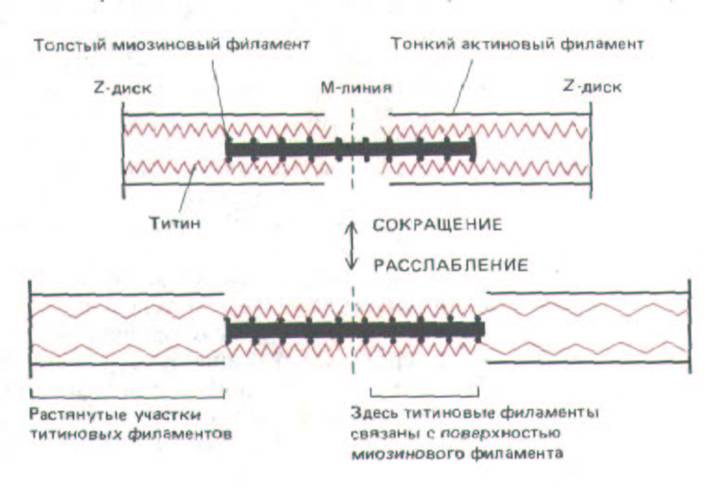
Рис. 11-21. Сеть из нитей титина, которые, как предполагается, соединяют в саркомерах скелетных мышц толстые миозиновые филаменты с Z-дисками. Эластичные титиновые нити, по-видимому, присоединены к толстым филаментам вдоль всей их поверхности, так что свободно изменять длину и обусловливать эластичность саркомера может только отрезок нити между концом толстого филамента и Z-диском.
Такая упругая сеть удерживает толстые филаменты точно посередине между Z-дисками и позволяет мышцам растягиваться за пределы области перекрывания толстых и тонких филаментов без разрушения саркомера. Поперечные мостики между толстыми и тонкими филаментами для простоты не показаны. (По К. Wang, J. Wright, J. Cell Biol. 107: 2199-2212, 1988.)
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
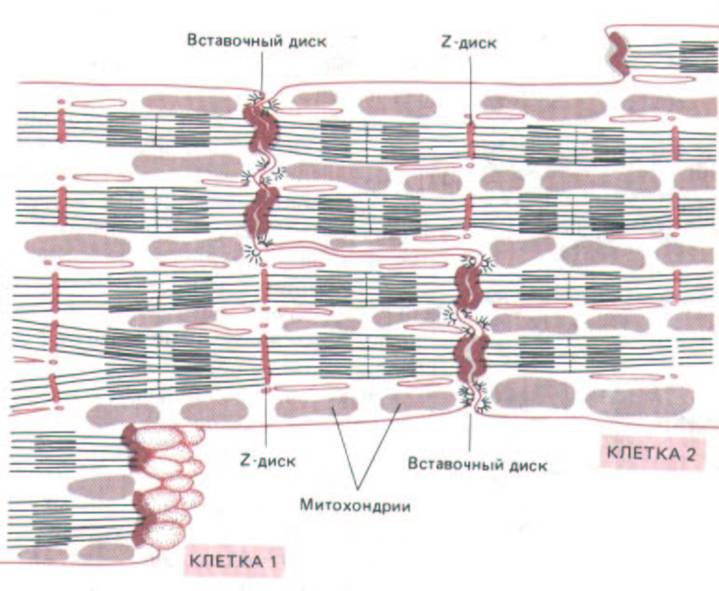
Рис. 11-22. Структура сердечной мышцы. Сердечная мышца состоит из множества отдельных клеток, каждая со своим ядром. Эти клетки соединены между собой с помощью специальных контактов, называемых вставочными дисками. В зоне каждого вставочного диска актиновые филаменты саркомеров соседних клеток входят в плотное вещество, связанное с плазматической мембраной, как если бы это был Z-диск. Таким образом, миофибриллы тянутся в мышце, игнорируя границы клеток.
мых клеток - миоэпителиалъных клетках (см. разд. 17,6)-работает механизм скользящих нитей.
Подобно скелетным мышцам, сердечная мышца выглядит исчерченной (поперечнополосатой), что отражает весьма сходную организацию актиновых и миозиновых филаментов. Сокращение тоже запускается сходным механизмом: потенциал действия, достигнув Т-
трубочек, вызывает выброс из саркоплазматического ретикулума Са2 +, который с помощью тропонин-тропомиозинового комплекса стимулирует сокращение. Однако клетки сердечной мышцы не являются многоядерными и соединены между собой конец в конец специальными вставочными
дисками (рис. 11-22). Вставочные диски выполняют по крайней мере три функции: 1) они соединяют при помощи десмосом (разд. 14.1.4) каждую клетку со следующей; 2) они связывают тонкие филаменты, входящие в состав миофибрилл соседних клеток (играя роль, аналогичную роли Z-
дисков внутри клетки); 3) в них находятся щелевые контакты (разд. 14.1.5), через которые потенциал действия быстро распространяется от клетки к клетке, синхронизируя их сокращение.
Наиболее «примитивная» мышца - в том смысле, что она имеет наибольшее сходство с немышечными клетками - лишена исчерченности, откуда ее название «гладкая» мышца. Она образует сократимые структуры желудка, кишки, матки, стенок артерий, железистых протоков и многих других частей тела, где необходимо медленное и продолжительное сокращение. Гладкомышечная ткань состоит из слоев удлиненных веретенообразных клеток, в каждой из которых одно ядро. В клетках есть и толстые, и тонкие филаменты, но они не организованы в столь упорядоченные структуры, как в скелетной мускулатуре и в сердце (в частности, они не образуют отдельных миофибрилл). Филаменты сократительного аппарата гладкомышечных клеток распределены более диффузно, хотя в основном они вытянуты вдоль длинной оси клетки, соприкасаясь под косым углом с плазматической мембраной в дисковидных контактах, соединяющих группы клеток вместе.
Сократительный аппарат гладкой мускулатуры неспособен к такому быстрому сокращению, как миофибриллы поперечнополосатых мышц. Однако он имеет то преимущество, что допускает гораздо большую степень укорочения и может поэтому осуществлять значительные пере-
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
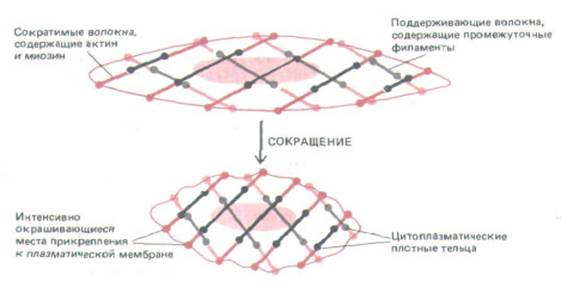
Рис. 11-23, Модель сократительного аппарата гладкомышечной клетки. На этой гипотетической схеме пучки сократительных филаментов, содержащих антин и миозин, присоединены одним концом к плазматической мембране, а другим концом - к несократимым пучкам промежуточных филаментов через цитоплазматические плотные тельца. Сократительные актомиозиновые пучки расположены с наклоном к длинной оси клетки (которая обычно гораздо сильнее вытянута, чем показано на схеме), и их сокращение намного укорачивает клетку. Показана только небольшая часть всех пучков.
мещения, несмотря на отсутствие такой системы рычагов, как кости. Благодаря какой организации актиновых филаментов и миозина это становится возможным, пока не ясно; одна из гипотетических моделей представлена на рис. 11-23.
|
|
|
|
|
Дата добавления: 2015-08-31; Просмотров: 2408; Нарушение авторских прав?; Мы поможем в написании вашей работы!