КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Транспозиционные взрывы приводят к существенным изменениям в геномах и повышают биологическое
10-36
10-36
10-36
10.5.8. Эволюция геномов ускоряется транспозирующимися элементами по крайней мере трех типов [60]
Геномы обычно содержат много разнообразных транспозирующихся элементов, или транспозовов. Впервые эти элементы были обнаружены в геноме кукурузы; некоторые из них удалось охарактеризовать и даже определить их первичную структуру. Лучше всего изучены транспозоны у дрозофилы, где известно более 30 их типов. Длина этих транспозонов колеблется от 2000 до 10000 нуклеотидных пар; большинство из них присутствует в геноме в количестве 5-10 копий на диплоидную клетку. В настоящее время различают три больших класса транспозонов на основании особенностей организации их последовательностей (табл. 10-3). Некоторые элементы перемещаются в геноме в виде ДНК, но есть и такие, у которых этот процесс включает образование промежуточного продукта (в его роли выступает РНК). В любом случае транспозоны способны размножаться, вырезаться из каких-то сайтов и внедряться в другие; их поведение можно охарактеризовать как паразитическое.
| Таблица 10-3. Три основных семейства транспозонов | |||
| Структура | Гены, входящие в состав полного элемента | Способ перемещения в геноме | Примеры |

| Кодируют транспозазу | Перемещаются в виде фрагмента ДНК или при вырезании, или при репликации | Р-элемент (дрозофила) Ac-Ds |
| (кукуруза) tn 3, IS1 (E.coli) | |||

| 3 (львиный зев) Tarn | ||

| Кодируют обратную транскриптазу, напоминают ретровирус | Перемешаются путем образования промежуточной формы РНК, синтезируемой с промотора, локализованного в LTR | Copia ТУ |

| ТНЕ-1 bsl | ||

| Кодируют обратную транскриптазу | Перемещаются путем образования промежуточной формы РНК, синтезируемой с соседнего промотора | Р-элемент (дрозофила) L1 (человек) |

| (кукуруза) cin 4 |
Длина этих элементов варьирует от 2000 до 12 000 нуклеотидных пар. В состав каждого семейства входит много различных элементов, в таблице приведены лишь немногие из них.
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
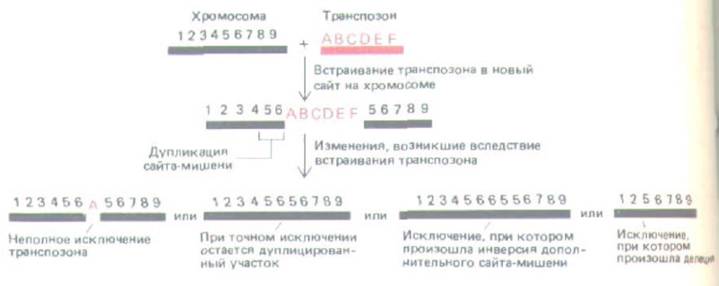
Рис. 10-70: Некоторые изменения в последовательностях ДНК хромосомы, возникающие вследствие перемещения транспозонов. При встраивании транспозона всегда образуется короткая дупликация хромосомной последовательности из 3-12 нуклеотидных пар. Ферменты сайт-
специфической рекомбинации, кодируемые этим элементом, участвуют и в последующем исключении транспозона. При таком исключении последовательность хромосомной ДНК часто не восстанавливается, как это показано на четырех примерах.
На долю транспоэонов приходится по крайней мере 10% геномнов ДНК высших эукариот. Большинство этих элементов перемещается лишь изредка, но поскольку их количество в клетке велико, транспозиция оказывает значительное влияние на разнообразие видов. Например, больше половины спонтанных мутаций, изученных у дрозофилы, вызвана встраиванием транспозона внутрь мутировавшего гена или вблизи него.
Мутации могут возникать либо когда элемент встраивается в ген, либо когда он начинает перемещаться в какое-либо другое место. Все известные транспозоны приводят к появлению коротких дупликаций в сайте-мишени, что связано с механизмом их встраивания (см. рис. 5-67, Б).
При вырезании транспозона из хромосомы обычно он оставляет на месте своего пребывания одну из копий, составляющих дупликацию (рис. 10-
70). Таким образом, перемещение транспозона сопровождается вставками и делециями в нуклеотидной последовательности.
Транспозоны вносят свой вклад в вариабельность генома и иными средствами. Если два транспозона, которые узнаются одним и тем же сайт-специфическим ферментом рекомбинации (транспозазоп), встраиваются в соседние сайты хромосомы, ДНК между ними может стать субстратом для транспозиции, осуществляемой с помощью транспозазы. Так как это весьма эффективный путь перемещения экзонов, справедливо утверждение, что транспозоны могут способствовать образованию новых генов (рис. 10-71).
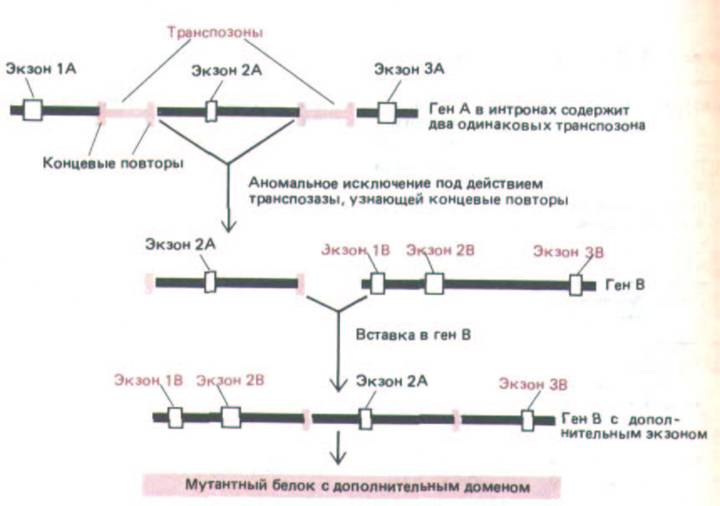
Рис. 10-71. Перемещение экзона, которое может происходить в результате встраивания транспозонов. Когда два транспозона, принадлежащие к одному и тому же типу (выделены цветом), оказываются по соседству друг с другом на хромосоме, то в транспозиции могут оказаться задействованными концы двух разных элементов (вместо двух концов одного и того же элемента); в результате хромосомная ДНК, заключенная между ними, переместится в новую область хромосомы. Так как по сравнению с экзонами интроны очень велики (см. рис. 9-7), изображенное на схеме встраивание нового экзона в ранее существовавший интрон - не такое уж невероятное событие.
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
10.5.9. Транспозоны могут влиять на регуляцию генов [61]
Перестройки последовательностей ДНК, вызываемые транспозонами, часто изменяют экспрессию близлежащих генов, что может привести к различным нарушениям в развитии животных или растений, например их пигментации (рис. 10-72). Большая часть таких изменений в регуляции генов, как правило, оказывается вредной для организма, но некоторые - могут оказаться и полезными.
Свойства мутаций, вызываемых транспозонами, необычны и позволяют отличить их от мутаций, возникших вследствие ошибок в репликации или репарации ДНК. Одно важное отличие состоит в том, что при перемещении транспозона вблизи гена часто оказываются новые последовательности, которые действуют как участки узнавания для сайт-специфических ДНК-связывающих белков, включая транспозазу
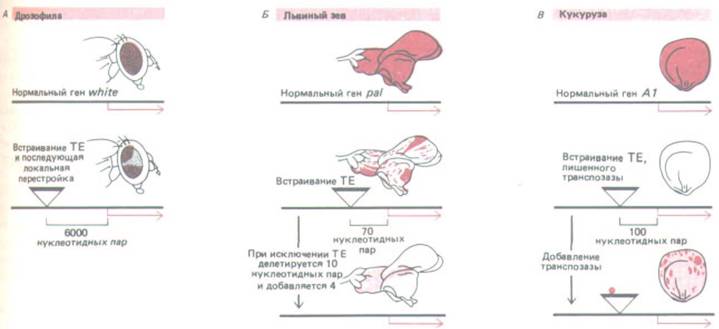
Рис. 10-72. Транспозоны могут обусловливать значительные изменения в регуляции генов. Для каждого из трех организмов приведен пример наследуемых изменений в распределении пигмента, вызванных встраиванием транспозонов (ТЕ) в регуляторные области генов.
Аналогичные процессы могут приводить к морфологическим изменениям в организме, воздействуя на рост и дифференцировку клеток. А.
Встраивание в регуляторные элементы, расположенные перед промотором гена white у дрозофилы, приводит к тому, что красный пигмент проявляется лишь в дорсальной и вентральной области глаза. Б. Встраивание перед промотором гена, определяющего образование пигмента у львиного зева, приводит к появлению цветов, у которых пегмент отсутствует везде, за исключением тех групп клеток, в которых этот элемент
был удален при транспози ции. Последующее исключение транспозона из генома всего растения обусловливает появление на ограниченном участке цветка бледной окраски. В. Пример регулируемого изменения окраски зерна кукурузы, вызываемого транспозоном. В данном случае транспозон действует как регуляторный белок, отчасти восстанавливающий окраску во всех клетках зерна, которое в ином случае осталось бы неокрашенным.
Кроме того, транспозаза катализирует случайное исключение элемента, в результате чего образуются отдельные интенсивно окрашенные пятна. (А
-по G.M. Rubm et aL, Cold Spring Harbor Symp. Quant Biol. 50: 329-335 1985; Б-no E S Coen, R. Carpenter and C. Martin, Cell 47: 285-296,1986; B-no Zs.
Schwarz-Sommer et at, EMBO J. 6: 287-294, 1987.)
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
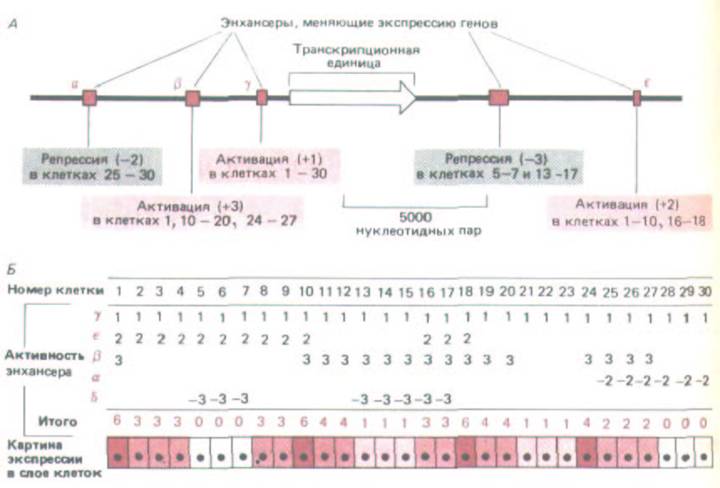
Рис. 10-73. Совместное действие отдельных энхансерных модулей приводит к характерному для каждой клетки порядку экспрессии генов. В связи с тем, что смесь регуляторных белков, связывающихся с каждым энхансером, варьирует от клетки к клетке, действие энхансера в различных клетках различно. Эта схема составлена по результатам, полученным на дрозофиле, где можно проанализировать много энхансеров, по-
отдельности выявляя их действие в трансгенных мухах. Для простоты стимулирующее (+) и ингибируюшее (—) действие каждого энхансера (α, β, γ, δ, или ε) оценивают числами от + 3 до —3 и считают, что эти числа можно складывать и вычислять суммарную активность энхансеров, определяющих уровень экспрессии генов.
и белки, регулирующие транскрипцию ДНК, входящей в состав транс-позона. Таким образом, эти последовательности могут действовать как энхансеры и усиливать транскрипцию генов, расположенных от них на расстоянии тысяч нуклеотидных пар. Пример такого типа воздействия на экспрессию гена пигмента у кукурузы приведен на рис. 10-72. Аналогичный эффект может вносить свой вклад в возникновение раковых клеток:
перенос регуляторных последовательностей в области, соседствующие с протоонкогеном, способен превратить его в онкоген.
Геномы высших эукариот, где длинные последовательности некодирующей ДНК перемежаются относительно короткими кодирующими участками, представляют собой «благодатную почву» для интеграции и исключения мобильных элементов. В связи с тем, что на транскрипцию генов влияют и удаленные от них на десятки тысяч нуклеотидных пар участки, можно ожидать, что многие возникшие при транспозиции изменения генома окажут влияние и на экспрессию генов. И напротив, по-видимому, лишь немногие перестройки приведут к разрушению коротких экзонов, содержащих кодирующие последовательности.
Может ли большой избыток некодирующей ДНК высших эукариот поддерживаться в ходе эволюции благодаря той регуляторной пластичности, которую она сообщает организму, содержащему много разнообразных транспозонов? То, что известно о регуляторных системах, контролирующих гены высших эукариот, находится в соответствии с этой возможностью. Энхансеры подобно экзонам, по-видимому, действуют как отдельные модули, и активность генов зависит от суммарного влияния на промотор набора энхансеров (рис. 10-73). Транспозоны, перемещая энхансеры по геному, могут способствовать оптимизации регуляции генов в целях выживания организма в ряду поколений.
разнообразие [62]
Другой уникальной особенностью, отличающей транспозоны от обычных мутагенов, является их способность долгое время находиться в состоянии покоя в хромосоме. Время от времени у части популяции бурно активируются движения транспозонов и, соответственно, их мутагенная
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
активность. При таких катастрофических изменениях в геномах, называемых транспозиционными взрывами, происходит почти одновременное перемещение транспозонов нескольких типов. Впервые транспозиционные взрывы были обнаружены в развивающихся растениях кукурузы.
Подобное явление наблюдается и при скрещивании определенных линий мух-феномен, известный под названием «гибридный дисгенез». Если такие взрывы происходят в клетках зародышевой линии, то они вызывают множественные изменения в геноме потомства отдельной мухи или растения.
Меняя свойства организма, транспозиционные взрывы повышают вероятность того, что два новых признака, каждый из которых сам по себе не обладает селективными преимуществами, окажутся выгодными, проявляясь вместе у единственной особи в популяции. Есть данные о том, что у определенных растений транспозиционные взрывы могут активироваться сильными стрессами, вызванными внешними условиями. Это приводит к появлению разнообразных, случайным образом модифицированных потомков, часть из которых может оказаться лучше приспособленной к выживанию в новых условиях. Возможно, что, по крайней мере у этих растений, существует механизм активации транспозонов, которые работают как мутагены и вызывают появление большого числа вариантных организмов в тот момент, когда такое разнообразие необходимо как никогда. Таким образом, транспозоны нельзя представлять себе только как паразитов; в некоторых случаях они способны действовать как полезные симбионты, способствуя выживанию тех видов, в геноме которых они содержатся.
|
|
Дата добавления: 2015-08-31; Просмотров: 1365; Нарушение авторских прав?; Мы поможем в написании вашей работы!