КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Точковые мутации обусловливают небольшие изменения генома, а его перестройка или увеличение
|
|
|
|
10-34
10-32
10-28
10-27
10-26
5-8
10-25
10.4.10. Многие мРНК-объект контроля на уровне трансляции [46]
Насколько контроль на уровне трансляции распространен у высших эукариот? Согласно некоторым оценкам, таким способом регулируется экспрессия одного гена из десяти. Контроль на уровне трансляции дает возможность клетке быстро и обратимо менять концентрацию белка, не подавляя синтез кодирующей его мРНК (см. разд. 12.4.7). По-видимому, экспрессия некоторых протоонкогенов регулируется именно так.
Особенно важную роль контроль на уровне трансляции играет в оплодотворенных яйцеклетках, где необходимо переключить синтез с белков, присущих ооциту в состоянии покоя, на белки, участвующие в быстром делении клеток. В таких яйцеклетках имеется большой запас мРНК, образовавшейся в ходе созревания ооцита. Многие из этих материнских мРНК до оплодотворения яйцеклетки не транслируются. Изучение яйцеклеток двустворчатых моллюсков показало, что до и после оплодотворения с рибосомами ассоциированы различные мРНК. При трансляции таких мРНК в бесклеточной системе образуются белки, соответствующие определенной стадии созревания яйцеклетки моллюска, однако это происходит лишь тогда, когда сохраняется мРНК в виде рибонуклеопротеина. Если РНК очистить от связанных с ней белков и затем транслировать, разница между белками, синтезированными на мРНК, взятой на разных стадиях, исчезает. Следовательно, фактор, определяющий, будет ли данная мРНК транскрибироваться, должен зависеть от того, как РНК связывается с регуляторними молекулами.
10.4.11. Сдвиг рамки трансляции приводит к образованию двух белков на одной молекуле мРНК [47]
Контроль на уровне трансляции, описанный выше, влияет на скорость инициации новых полипептидных цепей на молекуле мРНК.
Обычно
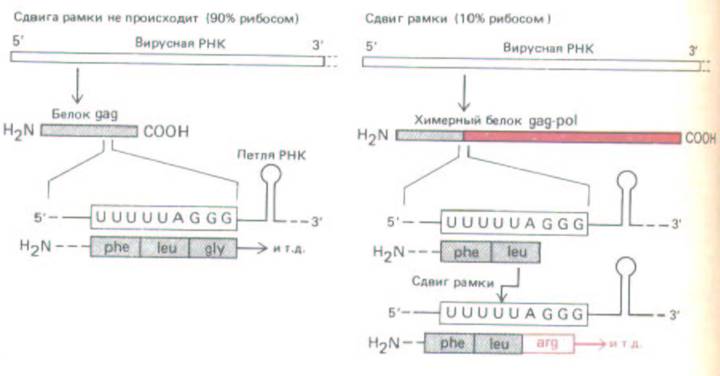
Рис. 10-58. Сдвиг рамки при трансляции необходим для образования обратной транскриптазы ретровируса. Вирусные обратная транскриптаза и интеграза образуются при расщеплении большого химерного белка gag-pol, а белки капсида в результате расщепления белка gag, присутствующего в больших количествах. Синтез обоих белков начинается в одной точке, но у gag он заканчивается на стоп-кодоне в той же рамке считывания, а при сдвиге рамки в направлении — 1 синтезируется химерный белок. Сдвиг рамки обусловлен локальными особенностями в структуре РНК (к ним относится и показанная на рисунке петля РНК), которые приводят к тому, что тРНКLeu, присоединенная к карбоксильному концу растущей полипептидной цепи, время от времени соскальзывает на один нуклеотид назад в рибосоме и спаривается с кодоном UUU вместо UUA, который определял ее включение. Представлена последовательность вируса иммунодефицита человека (ВИЧ-1). (По Т. Jacks et al., Nature 331: 280-283, 1988.)
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
завершение синтеза белка происходит автоматически. Однако в отдельных случаях образующийся белок может изменяться в ходе процесса, называемого сдвигом рамки трансляции (считывания).
Сдвиг рамки трансляции широко распространен у ретровирусов, что позволяет получать с одной мРНК различные количества двух или более белков. У этих вирусов обычно образуется большой полипротеин, который разрезается вирусной протеазой с образованием нескольких капсидных белков (gag-белки), обратной транскриптазы вируса и интегразы (роl-белки). Во многих случаях гены gag и роl находятся в разных рамках считывания, и, таким образом, сдвиг рамки необходим для образования рol-белков, присутствующих в гораздо меньших количествах. Сдвиг рамки происходит по определенному кодону в мРНК, и для его осуществления необходимо присутствие специфических последовательностей, которые могут находиться и перед этим сайтом, и за ним (рис. 10-58).
10.4.12. Экспрессия генов может контролироваться изменением стабильности мРНК [48]
Большая часть мРНК в клетках бактерий весьма нестабильна и имеет время полужизни около 3 минут. Так как мРНК у бактерий быстро синтезируется и также быстро разрушается, бактерия в ответ на изменения окружающей среды может соответствующим образом регулировать экспрессию своих генов.
Рнс. 10-59. Особые сигнальные последовательности, ответственные за нестабильность (быструю деградацию) некоторых мРНК. А. Три мРНК, время полужизни которых сильно различается. Непрерывный распад молекул мРНК, кодирующих различные факторы роста, обусловливает быстрое изменение их концентрации в ответ на внеклеточные сигналы. Гистоны необходимы, главным образом, для формирования хроматина, образующегося в ходе синтеза ДНК. Значительные изменения в стабильности их мРНК ограничивают время синтеза пистонов S-фазой клеточного цикла. Б. За необычно быстрый распад мРНК ответственны особые последовательности на З'-нетранслируемом конце молекулы. Представленные результаты получены в экспериментах, когда указанные измененные последовательности РНК синтезировались в клетках с генов, модифицированных методами генной инженерии.
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
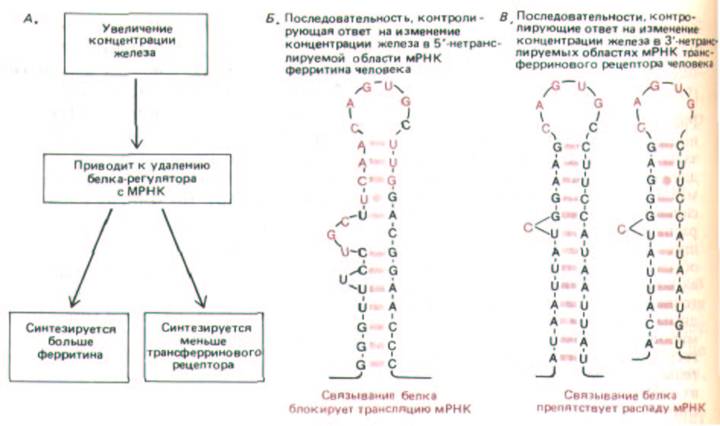
Рис. 10-60. В ответ на увеличение концентрации железа клетка повышает уровень синтеза ферритина (чтобы связать избыток железа) и снижает уровень синтеза рецепторов трансферрина (чтобы в клетку попадало меньше железа). Полагают, что оба типа ответов обусловлены одним и тем же регуляторным белком. Этот белок узнает общие свойства мРНК, кодирующих ферритин и рецептор трансферрина. А. Регуляторний белок отделяется от мРНК. Б. Элемент, контролирующий ответ на железо, расположенный в 5'-нетранслируемой области мРНК ферритина. Нуклеотиды, выделенные цветом, идентичны в мРНК ферритина млекопитающих, птиц и земноводных. Если поместить эту последовательность в 5'-
нетранслируемую область других мРНК, то их трансляция также будет контролироваться уровнем содержания железа в среде. В. Две из пяти одинаковых последовательностей, от которых зависит ответ на изменение концентрации железа в среде, расположенные в З'-нетранслируемой области мРНК, кодирующей рецептор трансферрина. Железо-зависимый контроль стабильности мРНК требует присутствия многих сайтов связывания регуляторных белков. Считается, что нуклеотиды, выделенные цветом, а также спиральный стебель шпильки, последовательность которого не так важна, играют решающую роль в узнавании белка. Поскольку контроль синтеза рецептора трансферрина и ферритина осуществляется разными механизмами, их уровни отвечают на изменения концентрации железа противоположным образом, несмотря на то, что в регуляции принимает участие один и тот же белок. (По М. W. Hentze et al., Science 238: 1570-1573, 1987; J. L. Casey et al., Science 240: 924-928, 1988.)
В эукариотических клетках мРНК более стабильна. Наприме мРНК, кодирующая 3-глобин, имеет время полужизни более 10 ч, есть и такие мРНК, время полужизни которых составляет всего 30 минут! или еще меньше. Нестабильные мРНК часто кодируют белки-регуляторы, концентрация которых в клетке быстро меняется (например, факторы роста и продукты протоонкогенов fоs и туc). Нетранслируемая З'-область многих таких нестабильных мРНК содержит длинные последовательности, обогащенные А и U, которые, по-видимому, и ответственны за их нестабильность (рис. 10-59).
Стабильность мРНК может меняться в ответ на внеклеточные сигналы. Так, например, стероидные гормоны действуют на клетку, не только усиливая транскрипцию отдельных генов (см. разд. 12.2.1), но и повышая стабильность некоторых мРНК, считываемых с этих генов. И наоборот, добавление к клеткам железа уменьшает стабильность мРНК, кодирующей трансферриновый рецептор, что приводит к образованию меньшего количества белка, связывающего железо. Интересно, что в дестабилизации мРНК трансферринового рецептора, по-видимому, участвует тот же чувствительный к железу белок, связывающийся с РНК, который контролирует трансляцию мРНК ферритина. В данном примере с трансферриновым рецептором белок соединяется с противоположным концом мРНК (нетранслируемый З'-конец), что обусловливает усиление, а не ослабление синтеза белка (рис. 10-60).
10.4.13. Для избирательной деградации мРНК необходим постоянный синтез белка [49]
Контроль стабильности мРНК в клетках эукариот лучше всего изучен для мРНК, кодирующих гистоны. Время полужизни этих мРНК во время S-фазы клеточного цикла (когда требуются новые гистоны) составляет около 1 ч, но при остановке синтеза ДНК сокращается до нескольких минут. Если в ходе S-фазы синтез ДНК подавить антибиотиками, гистоновая мРНК становится нестабильной; возможно, накопление свободных гистонов в отсутствие новой ДНК, с которой они могли бы связаться, специфически повышает скорость распада гистоновой мРНК.
Регуляция стабильности гистоновой мРНК зависит от короткой структуры, состоящей из петли и двухцепочечного участка на З'-конце;
она заменяет ро.уА-последовательность, присутствующую на З'-конце
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
других мРНК. Такой З'-конец образуется после того, как гистоновая мРНК синтезируется РНК-полимеразой II, в ходе специальной реакции расщепления, для которой необходимо спаривание с основаниями небольшой РНК, входящей в состав рибонуклеопротеиновой частицы (U7-мя РНК). Если методами генной инженерии соединить этот З'-конец с другими мРНК, они также становятся нестабильными при остановке синтеза ДНК (см. рис. 10-59). Таким образом, как и в случае мРНК других типов, скорость распада жестко контролируется сигналами, расположенными вблизи З'-конца, с которого, как полагают, и начинается деградация мРНК.
Если в середину кодирующей последовательности гистоновой мРНК поместить стоп-кодон, такая мРНК теряет способность быстро деградировать. Исходя из этого, было высказано предположение, что нуклеаза, ответственная за деградацию мРНК, соединена с рибосомой, и перед тем, как она начинает реакцию, большая часть гистоновой мРНК должна транслироваться. Это предположение могло бы объяснить, почему значительная часть нестабильных мРНК селективно стабилизируется при обработке клеток ингибитором белкового синтеза циклогексимидом. Тем не менее остается неясным, почему система деградации мРНК должна быть связана таким образом с рибосомой.
10.4.14. Некоторые мРНК расположены в определенных областях цитоплазмы [50]
Метод гибридизации in situ позволяет локализовать в клетке специфические молекулы мРНК (см. разд. 4.6.11). В гл. 16 будет идти речь о том, что некоторые мРНК, участвующие в формировании частей тела на ранних стадиях развития, сконцентрированы в определенных участках цитоплазмы ооцита (см. рис. 16-22, 16-60). Хотя остается неизвестным, с чем они при этом связаны, в ряде случаев подобное распределение по областям зависит от длинного нетранслируемого З'-конца мРНК.
Легче всего определить локализацию молекул мРНК в больших по размеру ооцитах, однако примеры специфической локализации мРНК известны и для соматических клеток. Вероятно, в будущем с помощью методов генной инженерии станет возможным менять расположение конкретных молекул мРНК в цитоплазме с тем, чтобы выявить эффект такого перемещения на функцию этих мРНК.
10.4.15. Редактирование мРНК изменяет смысл информационной РНК [51]
Открытие все новых молекулярных механизмов, используемых клеткой, заставляет биологов постоянно удивляться. Например, совсем недавно было обнаружено, что все мРНК у трипаносомы содержат общую кэпированную последовательность на 5'-конце, которая транскрибируется отдельно и затем добавляется к 5'-концам транскриптов гяРНК при сплайсинге двух ранее не связанных между собой молекул.
Подобный транссплайсинг имеет место у нематод, когда к ряду мРНК добавляется лидерная 5'-последовательность. Обнаружен он и у растений при комбинировании отдельных транскриптов РНК, из которых образуется кодирующая последовательность некоторых белков в хлоропластах.
Разрезание и воссоединение транскриптов может оказаться кратчайшим путем к появлению новых белков, а те немногие известные сегодня случаи такого соединения экзонов могут быть остатками процессов, некогда имевших гораздо более широкое распространение.
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
Другой любопытный феномен, в значительной мере изменяющий транскрипты РНК, был обнаружен в митохондриях трипаносом. Он заключается в редактировании РНК: в определенные области транскрипта добавляется или удаляется один (или более) нуклеотид U, что приводит к изменению исходной рамки считывания и меняет смысл информации. Однако неизвестно, как контролируется редактирование, в результате которого образуется последовательность, кодирующая необходимый белок.
Редактирование РНК, хотя и в более ограниченных пределах, вcтpeчается у млекопитающих, где с помощью этого механизма ген аполипопротеина В образует два типа транскриптов: в одном из транскритов цитозин, кодируемый ДНК, заменяется на урацил, в результате чего появляется стоп-кодон и синтезируется короткий тканеспецифичный вариант этого большого белка. Хотя до сих пор приведенный пример редактирования РНК остается единственным, представляется маловероятным, что этот феномен у млекопитающих ограничивается только одним геном.
10.4.16. Реакции, катализируемые РНК, вероятно, | имеют весьма древнее происхождение [52]
Все обсуждаемые в данном разделе механизмы контроля на посттранскрипционном уровне зависят от специфического узнавания определенной молекулы РНК; именно таким путем отбираются молекулы для специальных превращений, например, сплайсинга или деградация.
Узнавание возможно благодаря существованию большого числа сайт-специфических РНК-связывающих молекул, большая часть которых еще не охарактеризована. Сайты РНК, с которыми ассоциируют эти молекулы, обычно содержат группу открытых нуклеотидов в одноцепочечном участке (см. рис. 10-60). Следовательно, подобное сайт-специфическое связывание отличается от связывания с ДНК, где нуклеотидная последовательность обычно распознается по спаренным основаниям в двойной спирали. Более того, все известные молекулы, связывающиеся с определенными последовательностями ДНК, представляют собой белки; между тем к объединению со специфическими последовательностями РНК способны и белки, и молекулы РНК, принцип узнаваний у которых частично основан на комплементарном спаривании оснований РНК-РНК. Таким образом, пытаясь разобраться в посттранскрипционных процессах, мы вступили в мир РНК. Реакции, катализируемые молекулами РНК, гораздо труднее поддаются изучению, чем реакции, катализируемые белками. Большие молекулы РНК при выделении, из-за присутствия следовых количеств рибонуклеаз, легко распадаются, и их трудно очистить до гомогенного состояния, сохранив при этом, активность. Однако в настоящее врем»
методы генной инженерии с использованием очищенных РНК-полимераз позволяют получать in vitro большое количество чистых РНК с любой последовательностью (рис. 9-81). Благодаря этому появилась возможность подробно изучить химию катализируемых РНК реакций самосплайсинга (см. разд. 3.2.11) и определить минимальные размеры последовательностей, необходимых для саморасщепления РНК вироида растений (рис. 10-61).
Другой реакцией, в которой РНК играет роль катализатора как у прокариот, так и у эукариот, является расщепление предшественников тРНК комплексом белок—РНК, известным под названием РНКаза Р. РНК, входящая в состав мяРНП-частиц сплайсосом, вероятно, может также формировать и разрушать ковалентные связи (хотя это еще и не доказано). Полагают, что активный центр ферментативного комплекса пелтидилтрансферазы, участвующего в полимериза-
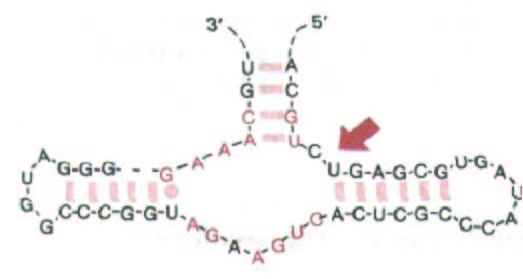
Рис. 10-61. Структура активного центра РНК вироида растений. Эта короткая молекула РНК сама расщепляет себя в участке, указанном стрелкой. Нуклеотиды, выделенные цветом, идентичны у семи саморасщепляющихся РНК, шесть из которых обнаружены в растениях и одна у животных. Реакции саморасщепления превращают тандемно повторенные последовательности одноцепочечной РНК (промежуточный продукт вироид-подобной РНК) в однокопийную линейную молекулу РНК, которая затем замыкается в кольцо. (По А. С. Forster et al., Nature 334: 265-267, 1988.)
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
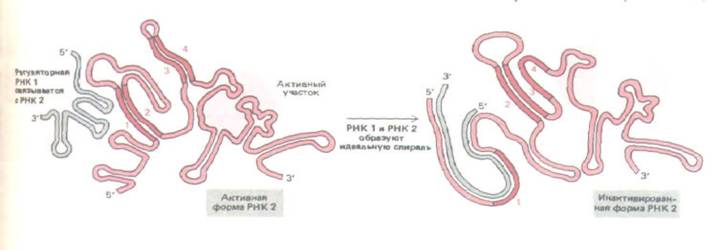
Рис. 10-62. Регуляторное взаимодействие двух молекул РНК способствует поддержанию постоянного числа копий в семействе бактериальных ДНК-содержащих плазмид ColEl. РНК (длиной около 100 нуклеотидов) представляет собой регуляторную молекулу, которая подавляет активность РНК 2 (длиной около 500 нуклеотидов), участвующей в инициации репликации плазмидной ДНК. РНК 1 комплементарна последовательности на 5'-конце РНК 2 и ее концентрация возрастает пропорционально числу молекул плазмидной ДНК в клетке. У РНК 2
последовательность 2 комплементарна как последовательности 1, так и последовательности 3 (ср. рис. 10-50) и способна замещаться при связывании РНК 1; в результате меняется конформация последовательности 4, инактивирующая РНК 2. (По Н. Masukata and J.Tomizawa, Cell 44:
125-136, 1986.)
ции аминокислот рибосомой, расположен на рРНК (см. разд. 5.1.8).
Молекулы РНК обладают также и регуляторной функцией. Антисмысловая РНК в клетках, измененных экспериментальным путем, делает эти клетки не способными экспрессировать определенный ген (механизм, аналогичный тому, который в норме регулирует экспрессию некоторых генов бактерий). Этот механизм на самом деле может иметь гораздо более широкое распространение. Особенно хорошо изученным примером такого рода служит контроль с обратной связью за началом репликации ДНК большого семейства бактериальных плазмид.
Контролирующая система ограничивает копийность плазмид, и таким образом не дает плазмидам убить хозяйскую клетку (рис. 10-62).
Изучение реакций, катализируемых РНК, представляет особый интерес для понимания хода эволюции. Как уже обсуждалось в гл. 1, первые клетки, по-видимому, не содержали ДНК, и в них было очень мало, а может быть и вообще не было белков. Многие из реакций, катализируемых РНК в современных клетках, могут представлять собой молекулярные ископаемые, т.е. происходить от той сложной сети реакций, направляемых РНК, которые предположительно преобладали в метаболизме клетки 3,5 млрд лет назад. Разобравшись в них, биологи, возможно, смогут проследить пути возникновения первой живой клетки.
Заключение
При осуществлении контроля экспрессии генов клетки воздействуют на многие стадии перехода РНК>белок. Полагают, что
большинство генов регулируется на нескольких уровнях, хотя преобладающим считают контроль на уровне инициации транскрипции. Тем не
менее, некоторые гены транскрибируются с постоянной скоростью, а их включение и выключение происходит только за счет воздействия на
РНК. К таким посттранскрипционным регуляторним процессам относят: 1) аттенуа-цию транскрипции путем преждевременной терминации, 2) альтернативный выбор сайта сплайсинга, 3) контроль за образованием З'-конца путем расщепления и добавления polyA, 4) контроль инициации
трансляции и 5) регулируемую деградацию мРНК. Для большинства из этих контролирующих процессов необходимо узнавание
последовательностей или структур в молекуле РНК. Такое узнавание может осуществляться либо белком-регулятором, либо регуляторной
молекулой РНК.
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
10.5. Организация и эволюция ядерного генома [53]
В геномах ныне живущих организмов записана значительная часть их эволюционной истории. Некоторые ее страницы могут быть расшифрованы при изучении последовательности ДНК этих организмов. Методы секвенирования ДНК, широко распространенные в настоящее время, дают возможность анализировать большое количество молекул ДНК и судить о том, как за десятки миллионов лет возникли гены, кодирующие определенные белки. Изучение случайных изменений, происходящих в хромосомах сейчас, проливает дополнительный свет на механизмы, ответственные за эволюционные изменения в прошлом. В данном разделе представлены некоторые молекулярно-генетические подходы, направленные на изучение организации и эволюции ядерного генома высших эукариот.
осуществляются в ходе генетической рекомбинации [54]
Последовательность нуклеотидов в ДНК должна точно реплицироваться и сохраняться. В гл. 5 обсуждались сложные механизмы, позволяющие ДНК наследоваться с необычайной точностью: за каждые 200000 лет случайно меняется лишь одна нуклеотидная пара из тысячи (см.
разд. 5.2). И даже при такой скорости мутирования в популяции, состоящей из 10000 особей, каждая возможная нуклеотидная замена будет «испробована» около пятидесяти раз за миллион лет. Если какой-либо вариант последовательности обладает преимуществом, он быстро размножится благодаря естественному отбору. Следовательно, можно ожидать, что у любого вида функция большинства генов будет оптимизирована в отношении вариаций, возникающих вследствие точковых мутаций.
Точковые мутации служат для тонкой «подстройки» генома, но долговременный эволюционный процесс должен быть связан с более радикальными генетическими изменениями. Эту функцию выполняет генетическая рекомбинация; с ее помощью геном может увеличиваться или уменьшаться (при дупликации или делеции), а его части могут перемещаться из одной области в другую, образуя новые комбинации.
Составляющие части генов (их экзоны и регуляторные элементы) могут перемешиваться, давая начало новым белкам, обладающим совершенно новыми функциями. Кроме того, если какой-либо ген представлен в геноме двумя копиями, одна из них может подвергнуться мутации, что приведет к дивергенции копий и их специализации для едва различающихся функций. Таким путем геном как целое постепенно усложняется и совершенствуется. Например, у млекопитающих почти каждый ген существует в нескольких вариантах: разные гены актина - для различных типов сократительных клеток, разные гены родопсина - для восприятия различных цветов, разные гены коллагена - для различных типов соединительных тканей и так далее. Экспрессия каждого гена регулируется строго и специфически. Изучение последовательностей ДНК показывает, что многие гены, даже значительно отличающиеся друг от друга, могут иметь родственные модульные области. Так например, определенная часть генов родопсина имеет общего предшественника с рядом генов, кодирующих некоторые гормоны и рецепторы (см. разд. 12.3.13); эта общая последовательность, вероятно, присутствует и в других белках (см. разд. 3.3.8).
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
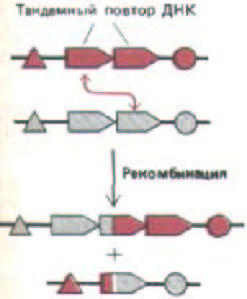
Рис. 10-63. Семейство тандемно повторенных генов теряет и восстанавливает свои копии вследствие кроссинговера между сестринскими хромосомами, несущими эти гены. Это происходит довольно часто, так как длинные участки гомологичных последовательностей ДНК являются хорошим субстратом для общей генетической рекомбинации.
Основой для возникновения подобных семейств генов и генных сегментов служит генетическая рекомбинация. Выше обсуждались молекулярные механизмы общей и сайт-специфической рекомбинации. В данном разделе будут рассмотрены некоторые результаты воздействия рекомбинации на геном.
|
|
|
|
|
Дата добавления: 2015-08-31; Просмотров: 575; Нарушение авторских прав?; Мы поможем в написании вашей работы!