КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
На примере семейства глобиновых генов можно проследить, как случайные дупликации ДНК способствуют
|
|
|
|
10-35
10-34
10-33
10,5.2. Тандемно повторяющиеся последовательности ДНК стремятся остаться неизмененными [55]
Дупликации генов обычно объясняют редкими событиями, которые катализируются некоторыми рекомбинационными ферментами.
Однако у высших эукариот имеется эффективная ферментативная система, которая соединяет концы разорванной молекулы ДНК. Таким образом, дупликации (а также инверсии, делеции и транслокации сегментов ДНК) могут возникать у этих организмов вследствие ошибочного воссоединения фрагментов хромосомы, которая по каким-то причинам оказалась разорванной. Если дуплицированные последовательности соединяются «голова к хвосту», то говорят о тандемных повторах. Появление одного тандемного повтора легко может привести к возникновению их длинной серии в результате неравного кроссинговера между двумя сестринскими хромосомами, поскольку длинные участки спаривающихся последовательностей представляют собой идеальный субстрат для обычной рекомбинации (рис. 10-63). Дупликация ДНК и следующий за ней неравный кроссинговер лежат в основе амплификации ДНК, процесса, который, как выяснилось, способствует возникновению раковых клеток (см. рис. 21-26). В ходе неравного кроссинговера число тандемно повторяющихся генов может как увеличиваться, так и уменьшаться (см. рис, 10-63). Большое количество повторяющихся генов будет поддерживаться естественным отбором лишь в том случае, если существование дополнительных копий окажется выгодным для организма. Как отмечалось выше, у позвоночных тандемный повтор кодирует большой предшественник рибосомной РНК, что необходимо для обеспечения потребности растущих клеток в новых рибосомах (см. разд. 9.4.16). Кластеры тандемно повторяющихся генов кодируют у позвоночных и другие структурные РНК, включая 5S-pPHK, U1- и U2-мяРНК. Тандемные повторы характерны и для гистоновых генов, на которых синтезируется большое количество белка, требующегося в каждой S-фазе.
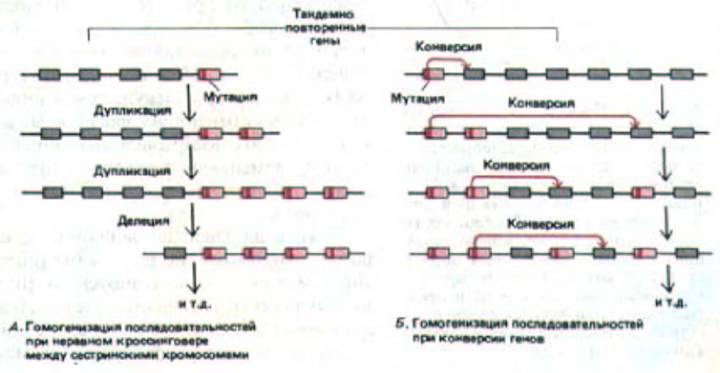
Рис. 10-64. Два типа событий, позволяющих сохранить последовательности ДНК в тандемном расположении и очень похожими друг на друга. А. Постоянное увеличение и уменьшение числа копий гена в тандеме при неравном кроссинговере (см. рис. 10-63) приводит к гомогенизации всех последовательностей генов, входящих в состав кластера. Б. При конверсии генов одна копия действует как матрица, которая передает либо все.
либо часть последовательностей своей ДНК другой копии гена. У высших эукариот эти процессы, по-видимому, присущи генам, расположенным рядом друг с другом на хромосоме. У низших эукариот, например у грибов, конверсия генов у которых изучена гораздо лучше, этот процесс, как оказалось, не лимитирован лишь соседними генами.
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
Можно ожидать, что в ходе эволюции последовательности тандемно расположенных генов, а также нетранскрибируемой ДНК спенсеров, расположенных между ними, дивергируют за счет случайных мутаций. изменяющих одну или несколько копий гена. Однако на самом деле последовательности тандемно повторенных генов и их спейсерная ДНК обычно почти идентичны. Полагают, что к этому причастны два механизма:
во-первых, неравный кроссинговер, приводящий к последовательному расширению и сокращению областей, содержащих тандемно повторяющиеся последовательности (анализ компьютерной модели такого кроссинговера показывает, что при этом последовательности имеют тенденцию оставаться прежними, рис. 10-64, А); во-вторых, конверсия генов (показано, что она может обусловливать гомогенизацию родственных последовательностей ДНК, рис. 10-64, Б).
эволюции организмов [56]
Дупликации ДНК имеют очень большое значение для эволюции новых белков. Чтобы убедиться в этом, обратимся к семейству глобиновых генов, поскольку его история изучена особенно хорошо. Явные гомологии в аминокислотной последовательности и в структуре современных глобиновых генов указывают на их происхождение от общего предка, несмотря на то, что некоторые члены этого семейства теперь расположены в геноме млекопитающих в совершенно разных местах.
Анализируя формы гемоглобина в организмах, стоящих на разных ступенях филогенетической лестницы, можно восстановить некоторые события, приведшие к возникновению разнообразных типов этого белка. Появление гемоглобиноподобных молекул в ходе эволюции, по-
видимому, способствовало увеличению размеров многоклеточных животных. Крупным животным для поддержания должного уровня кислорода в тканях уже недостаточно простой диффузии. В результате, гемоглобиновые молекулы обнаруживаются у всех позвоночных и многих беспозвоночных. Самая примитивная молекула, переносящая кислород, представляет собой глобиновую полипептидную цепь размером около 150
аминокислот. Она обнаруживается у многих морских червей, насекомых и примитивных рыб. Молекула гемоглобина у высших позвоночных устроена более сложно: в ее состав входит два типа глобиновых цепей. По-видимому, около 500 млн лет назад в ходе эволюции высших рыб произошла серия мутаций и дупликации соответствующего гена. В результате этих событий вначале образовалось два слегка отличающихся друг от друга гена, кодирующих цепи α- и β-глобинов в геноме каждой особи. У современных высших позвоночных каждая молекула гемоглобина представляет собой комплекс, состоящий из двух α- и двух β-цепей. (рис. 10-65). Такая структура функционирует гораздо более эффективно, чем молекула гемоглобина, содержащая одну цепь. Четыре кислород-связывающих сайта в молекуле α2β2 взаимодействуют друг с другом. Это взаимодействие приводит к кооперативному аллостерическому изменению в молекуле при связывании и освобождении кислорода, позволяющему доставлять в ткани гораздо большие порции кислорода.
В ходе дальнейшей эволюции млекопитающих мутации и дупликации, по-видимому, подвергся ген β-цепи, вследствие чего возник второй тип гемоглобина, синтезируемый только в эмбрионе. Образовавшаяся молекула гемоглобина обладает повышенным сродством к кислороду по сравнению с гемоглобином взрослой особи, и, таким образом, способствует переносу кислорода от матери к плоду. Ген, кодирующий
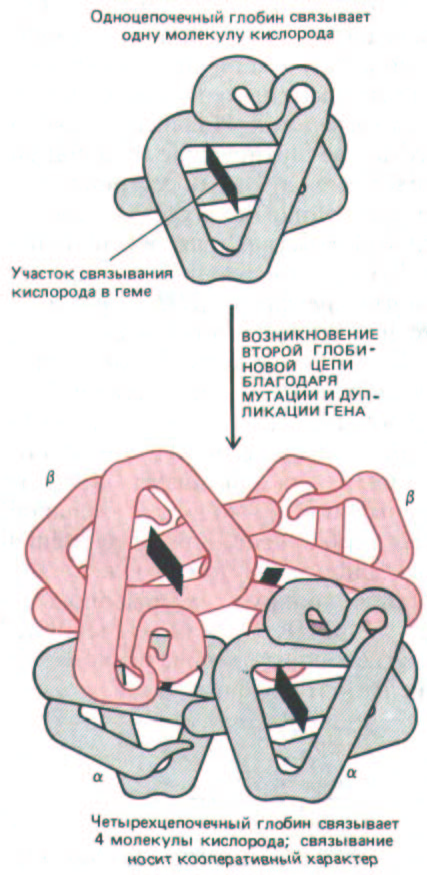
Рис. 10-65. Пространственная структура одноцепочечного и четырехцепочечного глобинов. Изображенный здесь четырехцепочечный гемоглобин представляет собой комплекс, состоящий из двух α- и двух β-глобиновых цепей. Глобин, состоящий из одной цепи, у некоторых примитивных позвоночных образует димер, который диссоциирует при связывании кислорода и представляет собой промежуточную ступень в эволюции глобина, содержащего четыре цепи.
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
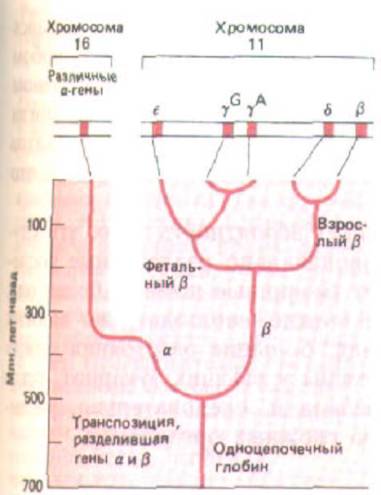
Рис. 10-66. Схема эволюции цепей глобина на примере семейства β-подобиых глобиновых генов (см. рис. 10-39). Относительно недавно возникшие дупликации гена γ -цепи, привели к образованию γG- и γА-цепей, которые являются (3-подобными и обладают идентичными функциями.

Рис. 10-67. Структура молекулы антитела (иммуноглобулинов). Эта молекула состоит из двух идентичных тяжелых цепей и двух идентичных легких цепей (выделены цветом). Каждая тяжелая цепь содержит четыре сходных ковалентно связанных домена. Каждая легкая цепь имеет в своем составе два таких домена. Каждый домен кодируется отдельным экзоном; вероятно, все экзоны возникли в результате дупликации одного исходного экзона.
новую, похожую на β-цепь, молекулу гемоглобина, вновь подвергся последовательным мутациям и дупликациям, в ходе которых возникли два новых гена ε и γ. Цепь. синтезируется на более ранних стадиях развития (с образованием α2ε2), чем γ-цепь эмбриона, образующая форму α2γ2 (см.
рис. 10-39, Б). Дупликация гена β-цепи взрослых, происшедшая еще позже в ходе эволюции приматов, привела к образованию гена δ-глобина и соответственно минорной формы глобина (α2δ2), обнаруживаемой только у взрослых приматов (рис. 10-66). Каждый из этих дуплицировавшихся генов впоследствии был модифицирован за счет точковых мутаций, воздействующих на свойства конечной молекулы гемоглобина, а также в результате изменений в регуляторных областях, определяющих выбор времени и уровень экспрессии данного гена (см. рис. 10-73).
Конечный результат процесса дупликации генов, приведшего к дивергенции глобиновых цепей, хорошо виден при рассмотрении генов, возникших из исходного β-гена и расположенных в виде серии гомологичных последовательностей ДНК внутри сегмента ДНК размером 50000
нуклеотидных пар (см. рис. 10-39, А). У человека кластер α-глобиновых генов находится на другой хромосоме. На основании того, что у птиц и млекопитающих кластеры α- и β-глобиновых генов обнаруживаются в разных хромосомах, а у лягушки Xenopus они лежат рядом, считается, что два гена разъединились в результате транслокации примерно 300 млн лет назад (рис. 10-66). Подобные транслокации, вероятно, способствуют стабилизации дуплицированных генов, обладающих различными функциями, поскольку предохраняют их от гомогенизации, которой часто подвергаются близлежащие гены со сходной последовательностью (см. рис. 10-64).
Существует несколько дуплицированных последовательностей глобиновой ДНК, входящей в состав кластеров α- и β-глобиновых генов, которые не являются активными. Это пример псевдогенов, которые имеют высокую степень гомологии с активными генами, но неактивны вследствие мутаций, препятствующих их экспрессии. Существование подобных псевдогенов не должно вызывать удивления, ведь не все дупликации ДНК могут приводить к возникновению новых активных генов, между тем неактивные последовательности не удаляются из генома немедленно.
Сравнивая последовательности ДНК многих семейств генов у животных, стоящих на разных ступенях филогенеза, можно проследить значительную часть истории нашей эволюции (см. рис. 4-62).
10.5.4. Гены, кодирующие новые белки, могут образовываться при рекомбинации экзонов [54]
Роль дупликации ДНК в эволюции не ограничивается их участием в образовании больших генных семейств. Дупликации могут иметь значение и для возникновения новых одиночных генов. Белки, кодируемые такими генами, можно узнать по присутствию в них повторяющихся сходных белковых доменов, которые последовательно ковалентно связаны друг с другом. Например, иммуноглобулины (рис. 10-67), альбумины, а также большинство фибриллярных белков (таких, как спектрины и коллагены) кодируются генами, возникшими в результате многократных дупликации исходной последовательности ДНК.
У генов, возникших таким путем, каждый экзон часто кодирует отдельную субъединицу или домен в белке (см. разд. 3.3.4). Организация кодирующих последовательностей ДНК в виде серии таких экзонов, разделенных длинными интронами, в значительной мере упростила эволюцию новых белков. Например, дупликации, необходимые для
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
образования отдельного гена, кодирующего белок с повторяющимися доменами, могут возникать при разрыве и воссоединении ДНК в любом месте длинных интронов, окружающих экзон. Без интронов в исходном гене было бы лишь несколько сайтов, рекомбинация по которым могла бы привести к дупликации домена. Увеличивая число возможных сайтов для дупликации, интроны значительно повышают вероятность того, что дупликация окажется полезной.
Наличие интронов намного увеличивает вероятность того, что случайная рекомбинация соединит две первоначально разделенные последовательности ДНК, которые кодируют различные домены белка (см, рис. 10-71). Результаты таких событий можно наблюдать во многих современных белках (см. рис. 3-38). Итак, большие расстояния между экзонами, кодирующими отдельные домены у высших эукариот, ускоряют процесс возникновения новых белков и, следовательно, увеличивают эффективность эволюции весьма сложных организмов.
10.5.5. Вероятно, большинство белков кодируются генами, состоящими из многих небольших экзонов [57]
Открытие в 1977 г. прерывистости генов эукариот оказалось совершенно неожиданным. Все исследованные до этого гены были бактериального происхождения и не содержали интронов. У бактерий, как известно, отсутствуют ядро и внутренние мембраны, их геном меньше, чем геном эукариот, и традиционно считалось, что бактерия напоминает ту древнюю простую клетку, из которой произошла клетка эукариотическая. Неудивительно поэтому, что многие биологи вначале воспринимали интроны как причудливую позднейшую эволюционную добавку. Однако в настоящее время все больше утверждается точка зрения, согласно которой прерывистые гены имеют весьма древнее происхождение, а бактерии потеряли свои интроны лишь после того, как возникла большая часть их белков.
Мысль о том, что интроны появились в ходе эволюции очень давно соответствует современному представлению о происхождении белков методом проб и ошибок при рекомбинации отдельных экзонов, кодирующих различающиеся белковые домены. Более того, доказательства древнего происхождения интронов были получены при изучении генов, кодирующих распространенный фермент триозофосфатизомеразу.
Триозофосфатизомераза играет важную роль в метаболизме всех клеток, катализируя центральное событие при гликолизе и глюконеогенезе-
взаимопревращение глицералальдегида-3-фосфата и дигидроксиацетон-фосфата (см. рис. 2-38). Сравнивая аминокислотную последовательность этого фермента у различных организмов, можно сделать вывод, что фермент возник еще до дивергенции прокариот и эукариот от общего предка, поскольку 46% аминокислотной последовательности у человека и бактерии идентичны. У позвоночных (курицы и человека) ген, кодирующий этот фермент, содержит шесть интронов, причем пять из них присутствуют точно в том же месте у кукурузы. Из этого следует, что эти пять интронов существовали в гене до того, как растения и животные дивергировали в ходе эволюции эукариот, что, как установлено, произошло 109 лет назад (рис. 10-68).
Мелкие одноклеточные организмы находятся под сильным давлением отбора, что заставило их воспроизводиться путем деления клеток с максимальной скоростью, какую только позволяет содержание питательных веществ в окружающей среде. В связи с этим, они вынуждены свести к минимуму содержание ненужной ДНК, которую надо синтезировать в каждом цикле клеточного деления. Для организмов большего размера, живущих благодаря хищничеству, и в целом для многокле-
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
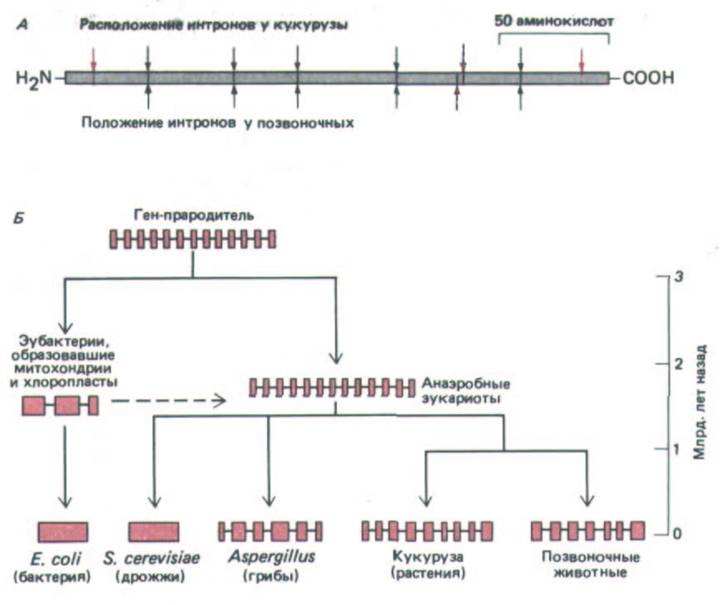
Рнс. 10-68. Эволюция прерывистых генов с древнейших времен. А. Структура гена триозофосфатизомеразы у растений и животных.
Одинаковые положения интронов у кукурузы (зерна) и позвоночных отмечены черными стрелками, а отличающиеся положения выделены красными стрелками. Так как считается, что растения и животные возникли от общего предка около миллиарда лет назад, общие интроны должны иметь очень древнее происхождение. Б, Гипотетическая схема возникновения определенного гена. Последовательности экзонов выделены цветом, а последовательности интрона обозначены черным. Приведенный здесь ген кодирует белок, необходимый для всех клеток. Подобно триозофосфатизомеразе, этот белок, по-видимому, сформировал свою окончательную трехмерную структуру перед тем, как от общего предка отделились бактерии, архебактерии и эукариоты. Этот общий предок обозначен на рисунке как «ген-прародитель». Пунктиром указано примерное время прохождения эндосимбиотических процессов, которые привели к возникновению митохондрий и хлоропластов. (А-по W. Gilbert, M.
Marchionni and G.Mc. Knight, Cell 46: 151-154, 1987.)
точных организмов, у которых скорости деления клеток определяются другими причинами, такое сильное давление отбора, ведущее к удалению избыточной ДНК из генома, отсутствует. Скорее всего именно эти обстоятельства объясняют, почему бактерии должны были потерять свои интроны, тогда как эукариоты их сохранили. Подобное объяснение соответствует также другим данным, полученным при изучении триозофосфатизомеразы: в то время как многоклеточный гриб Aspergillus содержит пять интронов в гене, кодирующем этот фермент, его одноклеточный родич, дрожжи Saccharomyces, не содержит их вообще.
Каков же механизм потери интронов? Возможно, интроны терялись при постепенных случайных делециях коротких сегментов ДНК, но более вероятно, что эукариотические клетки (а возможно, также и предки бактерий) имеют механизм точной и селективной делеции всего интрона из своих геномов. Например, в клетках большинства позвоночных содержится лишь один ген инсулина с двумя интронами, но у крыс no-соседству имеется еще один инсулиновый ген, в составе которого всего один интрон. Очевидно, второй ген возник относительно недавно в результате дупликации и затем потерял один из своих интронов. Так как при потере интрона необходимо точное воссоединение кодирующих последовательностей ДНК, считается, что второй ген возник в результате редкого события - включения в геном ДНК-копии мРНК соответствующего гена, откуда интроны были точно удалены. Подобные копии, не содержащие интронов, могут появляться благодаря активности обратных транскриптаз. Считают, что ферменты рекомбинации дают возможность таким копиям спариться с исходной последовательностью, которая затем «корректируется» по матрице, лишенной интронов, в ходе событий, напоминающих конверсию гена.
Обратные транскриптазы синтезируются в клетках транспозируемыми элементами (см. табл. 10-3) и всеми ретровирусами. Образование ДНК-копий частей генома при обратной транскрипции, по-видимому, тоже внесло свой вклад в эволюцию геномов высших организмов.
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
|
|
|
|
|
Дата добавления: 2015-08-31; Просмотров: 2133; Нарушение авторских прав?; Мы поможем в написании вашей работы!