КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Какой-то цитоплазматический сигнал задерживает подготовку к митозу, пока не завершена репликация
|
|
|
|
13-6
13-5
13-4
13-3
Фазы клеточного цикла и их причинные взаимосвязи
Под микроскопом деление эукариотической клетки выглядит как поразительный спектакль. При митозе содержимое ядра конденсируется,
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
образуя видимые хромосомы, которые в результате серии удивительно согласованных движений разделяются на два дочерних набора; затем при цитокинезе сама клетка делится на две дочерние, каждая из которых получает один из двух наборов хромосом. Благодаря легкости, с которой их можно наблюдать, митоз и цитокинез были в центре внимания ранних исследователей. Однако эти два события вместе составляют лишь короткую фазу всего клеточного цикла, известную как фаза М (от слова «митоз»). Гораздо более длительный период между двумя последовательными М-
фазами известен как интерфаза. Под микроскопом интерфаза обманчиво выглядит как антракт без всякого действия, когда клетка просто медленно увеличивается в размерах. Более изощренные методы позволяют узнать, что на самом деле интерфаза - это период, во время которого в строго заданной последовательности происходят сложные приготовления к митозу. В этом разделе мы обсудим, как можно изучать последовательность событий в интерфазе и каковы причинные взаимосвязи между фазами клеточного цикла.
13.1.1. Репликация ядерной ДНК происходит в определенный период, составляющий часть интерфазы [1]
В большинстве клеток синтез ядерной ДНК занимает лишь некоторую часть интерфазы - период, называемый S-фазой клеточного цикла.
Обычно между концом М-фазы и началом синтеза ДНК имеется интервал, известный как фаза G1 (от англ, gap-промежуток); другой интервал, называемый фазой G2, отделяет конец синтеза ДНК от начала следующей М-фазы. Таким образом, интерфаза состоит из последовательности фаз G1, S и G2 и обычно занимает не меньше 90% всего времени клеточного цикла. Например, в быстро делящихся клетках
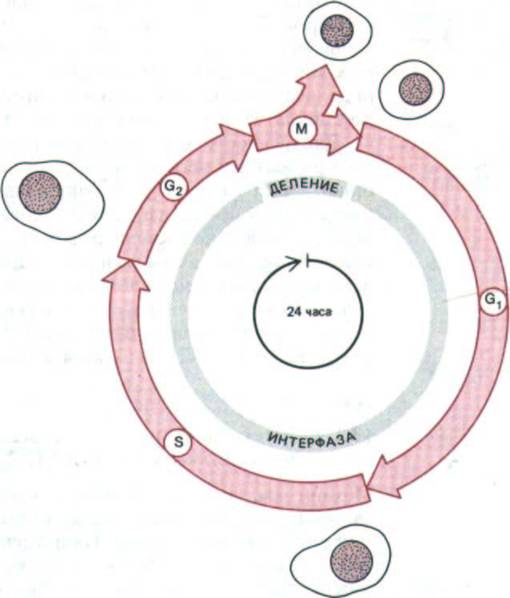
Рис. 13-1. Четыре последовательные фазы клеточного цикла типичной эукариотической клетки. После фазы М, которая состоит в делении ядра (митоз) и цитоплазмы (цитокинез), дочерние клетки вступают в интерфазу нового цикла. Интерфаза начинается с фазы G1, в которой возобновляются интенсивные биосинтетические процессы, резко замедленные во время митоза. Фаза S-это период синтеза ДНК; она заканчивается, когда содержание ДНК в ядре удвоится и хромосомы полностью реплицируются (теперь каждая хромосома состоит из двух идентичных «сестринских хроматид»). Затем клетка вступает в фазу С2, которая продолжается до начала митоза, т.е. фазы М. В фазе М удвоившиеся хромосомы конденсируются и становятся хорошо видимыми в световой микроскоп. Ядерная оболочка разрушается (исключение составляют одноклеточные эукариоты, например дрожжи, - у них она остается интактной); сестринские хроматиды расходятся и формируют два новых ядра, а цитоплазма делится с образованием двух дочерних клеток, имеющих по одному ядру. Процесс цитокинеза завершает фазу М, и начинается интерфаза следующего клеточного цикла. На рисунке представлен типичный 24-часовой цикл, однако длительность клеточного цикла у высших эукариот сильно варьирует: она может быть короче 8 часов, а у взрослых животных - больше года, причем различия в основном зависят от продолжительности фазы G1.
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
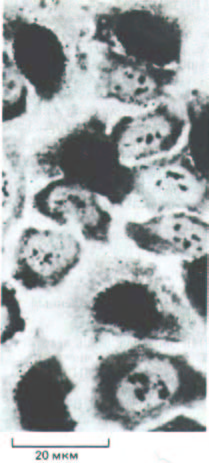
Рис. 13-2. Радиоавтограф, полученный после кратковременной инкубации клеток с 3Н-тимидином. Эта методика описана в разд. 4.5.2.
Наличие зерен серебра в фотоэмульсии над ядрами (черные участки) свидетельствует о том, что клетки включали 3Н-тимидин в ДНК и, следовательно, в период экспозиции какое-то время находились в фазе S. (С любезного разрешения James Cleaver.)
высших эукариот М-фаза обычно повторяется раз в каждые 16-24 ч, а сама длится только от 1 до 2 ч. Типичный клеточный цикл с его четырьмя последовательными фазами показан на рис. 13-1; некоторые подробности приведены в подписи к этому рисунку.
Время синтеза ДНК в клеточном цикле было впервые выявлено в начале 1950-х гг. с помощью метода радиоавтографии. В стандартном методе применяют 3Н-тимидин - радиоактивный предшественник соединения, которое клетка использует исключительно для синтеза ДНК, 3Н-
тимидин можно либо инъецировать животному для изучения циклов клеточного деления в тканях, либо добавлять в культуральную среду in vitro (рис. 13-2). В первом случае через определенное время после введения 3Н-тимидина у животного берут ткань и приготовляют из нее радиоавтографы. Те клетки, которые в прошедший период синтезировали ДНК (и, следовательно, находились в S-фазе), могут быть выявлены по зернам серебра над их ядрами. Подсчитывая долю клеток, находившихся в М-фазе, при различной длительности включения 3Н-ти-мидина, можно показать, что в клеточном цикле имеются четыре фазы, описанные выше, и измерить продолжительность каждой из них.
Предположим, что делают одну инъекцию и через короткое время, скажем через полчаса, ткань фиксируют для радиоавтографии. В типичной клеточной популяции, где все клетки делятся быстро, но не синхронно, около 30% клеток получат радиоактивную метку. Это будут те клетки, которые синтезировали ДНК в короткий период экспозиции в присутствии 3Н-тимидина, и их доля в клеточной популяции отражает долю клеточного цикла, занятую S-фазой (рис. 13-3). Только около 5% клеток в момент фиксации окажутся в стадии митоза (малая величина этого митотического индекса означает, что митоз занимает лишь небольшую часть клеточного цикла), но ни в одной из них не будет радиоактивной метки. Это указывает на то, что фазы М и S-обособленные части клеточного цикла. Однако, с другой стороны, если препараты фиксировать через несколько часов после введения 3Н-тимидина, то некоторые клетки, находящиеся в митозе, получат радиоактивную метку; вероятно, эти клетки еще синтезировали ДНК в момент инъекции. Минимальный интервал между инъекцией и временем появления меченых митотических клеток будет равен длительности фазы G2. Такого рода исследования позволяют определить длительность всех четырех фаз цикла. Пример использования этого метода приведен на рис. 13-4.
Длительность клеточных циклов в разных тканях, у разных видов и на разных стадиях очень широко варьирует - она может быть меньше одного часа (например, в раннем эмбрионе лягушки) и больше года (например, в печени взрослого человека). Хотя различаться по длительности в известной степени могут все фазы клеточного цикла, это в особенности касается фазы G1, длительность которой может варьировать в пределах практически от нуля (в раннем зародыше лягушки) до столь больших величин, что клетки кажутся вообще прекратившими деление (в зрелой печени человека). Часто говорят, что клетки в такой покоящейся фазе G1 находятся в состоянии G0 (см. разд. 13.3.8).
13.1.2. Клеточный цикл легче всего изучать на культурах in vitro [1, 2]
Механизмы, лежащие в основе клеточного цикла, трудно изучать в сложных и недоступных тканях интактного животного. Легче работать с клеточными культурами. Например, с помощью цейтраферной съемки (разд. 4.1.5) можно наблюдать, как отдельная клетка претерпевает митоз, растет, затем опять входит в стадию митоза; это делает воз-
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
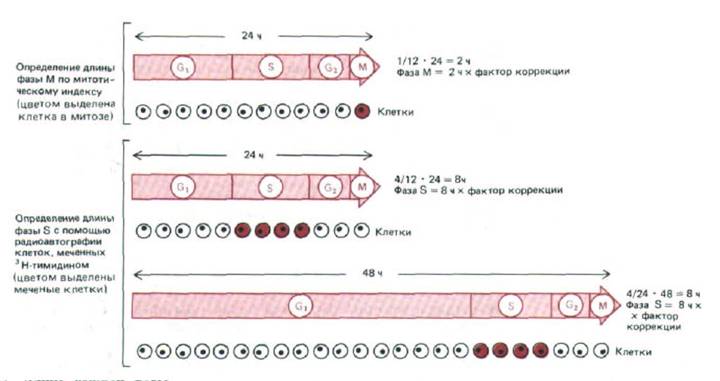
Рис. 13-3. Длина каждой фазы клеточного цикла примерно равна доле клеток, находящихся в этой фазе в каждый данный момент, умноженной на общую продолжительность цикла (если популяция клеток растет равномерно и все клетки делятся с одной и той же скоростью).
Однако для точного расчета длительности каждой фазы нужен «фактор коррекции», который изменяется в пределах от 0,7 для клеток ранней фазы G1 до 1,4 для митотических клеток. Этот коэффициент необходим потому, что в равномерно растущей популяции всегда больше молодых (недавно поделившихся) клеток, чем старых.
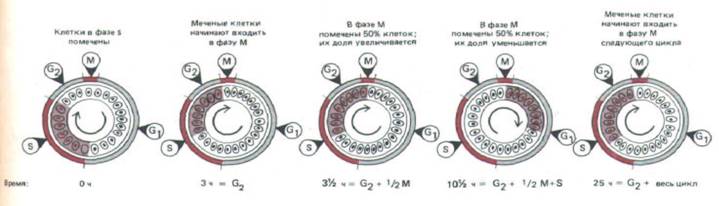
Рис. 13-4. Метод, используемый обычно для измерения продолжительности фаз клеточного цикла. Несинхронно пролиферирующую клеточную популяцию короткое время выдерживают в среде с 3Н-тимидином, а затем отмывают; при этом клетки продолжают проходить свой цикл. Через разное время после экспозиции берут пробы клеток и получают радиоавтографы. Для интерпретации результатов полезно изобразить клетки так, как будто они распределены на равномерно вращающемся круге, причем положение каждой клетки соответствует стадии цикла, на которой она находится. Вначале клетки в фазе S получают радиоактивную метку (окрашенный участок), а клетки в фазах G2, М и G1 ее не получают.
Через промежуток времени, равный длительности G2, меченые клетки начинают вступать в фазу М; когда пройдет время, равное G2 + М, получают метку все клетки, находящиеся в фазе М, и т.д. Отмечая время вступления меченых клеток в фазу М и последующие фазы вплоть до возвращения в М, в принципе можно определить среднюю длительность фаз G2, М и S и общую продолжительность цикла, а отсюда (путем вычитания) и длительность G,. В приведенном примере эти величины равны 3, 7, 22 и 11 ч соответственно.
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
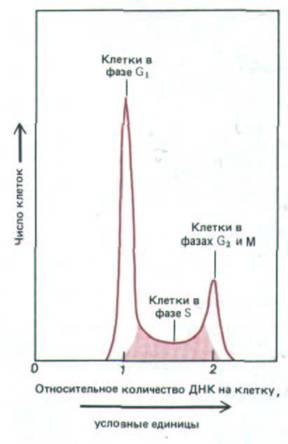
Рис. 13-5. Типичные данные, полученные при анализе содержания ДНК в отдельных клетках растущей популяции с помощью флуоресцентного клеточного анализатора - электронного прибора, основанного на том же принципе, что и сортировка флуоресцентно-окрашенных клеток (см. разд. 4.3.1). Клетки окрашивают красителем, приобретающим способность флуоресцировать при связывании с ДНК. Поэтому уровень флуоресценции прямо пропорционален содержанию ДНК в каждой клетке. Клетки подразделяются на три категории: с нереплицированной ДНК (1
условная единица), т.е. находящиеся в фазе G1, с полностью реплицированной ДНК (2 условные единицы), т. е. находящиеся в фазе G2 или М, и с промежуточным содержанием ДНК, находящиеся в фазе S. Распределение клеток в представленном примере показывает, что число клеток в фазе G1
больше, чем в G2 и М, взятых вместе. Это означает, что в данной популяции фаза G1 длиннее, чем G2 + M.
можным прямое измерение длительности М-фазы и всего клеточного цикла. Клетки, осуществляющие синтез ДНК в культуре, можно выявлять таким же образом, как в интактном организме, - методом радиоавтографии с использованием 3Н-тимидина. Можно также прослеживать ход клеточного цикла путем прямого измерения содержания ДНК в клетке; эту задачу сильно облегчает применение флуоресцентного анализатора
клеток (рис. 13-5).
Дальнейшее упрощение анализа клеточного цикла состоит в использовании большой популяции культивируемых клеток, одновременно проходящих одни и те же фазы клеточного цикла. Такие синхронные клеточные популяции можно получать разными способами. Самый ранний метод состоял в выдерживании клеток в растворе вещества, нарушающего определенную стадию клеточного цикла; длительное пребывание культуры в таком растворе приводит к тому, что все клетки останавливаются на этой стадии, а после снятия блокады возобновляют цикл и проходят его синхронно. Однако в ходе клеточного цикла параллельно осуществляется много различных процессов, и вряд ли все они будут блокироваться одновременно. Поэтому было бы лучше по возможности применять такие методы получения синхронных популяций, которые не нарушают нормальное прохождение клеточного цикла. Для большинства клеток млекопитающих самый простой и лучший

Рис. 13-6. Обычный метод получения синхронной популяции животных клеток в культуре. Митотические клетки отбирают, стряхивая их с поверхности чашки, в которой они растут. После перенесения в новую чашку эти клетки проходят дальнейшие циклы синхронно. Из-за случайных различий в скорости деления индивидуальных клеток синхронность после нескольких циклов деления теряется.
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
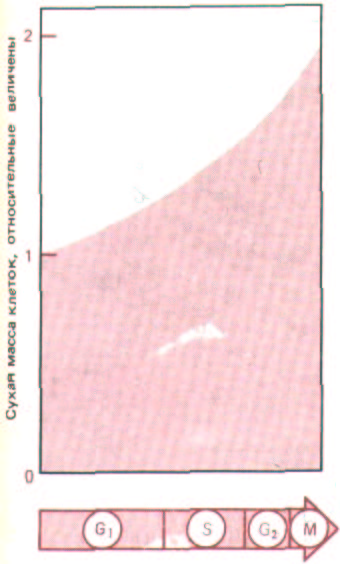
Рис. 13-7. График увеличения массы клеток за время клеточного цикла. Большая часть клеточных компонентов синтезируется более или менее равномерно на протяжении всей интерфазы, причем скорость их образования обычно возрастает по мере того, как размеры клетки и ее биосинтетическая активность увеличиваются (с коротким перерывом в фазе М). Чтобы в пролиферирующей популяции средняя величина клетки оставалась постоянной, количество каждого компонента за время цикла должно в точности удваиваться.
метод состоит в том, что используют изменения цитоскелета в М-фазе, приводящие к «округлению» клеток. Округлившиеся в М-фазе клетки так слабо прикрепляются к дну культуральной чашки, что их можно отделить легким встряхиванием (рис. 13-6). Митотические клетки, отобранные таким способом, составляют синхронную популяцию, в которой почти сразу же наступает фаза G1 клеточного цикла. Применяют и другой метод:
поскольку клетки по мере прохождения цикла увеличиваются в размерах, можно использовать центрифугирование, чтобы выделить субпопуляции клеток, находящихся на разных стадиях цикла.
13.1.3. Критические события клеточного цикла наступают внезапно на фоне непрерывного роста клеток [3]
На синхронной популяции делящихся клеток можно более детально изучать химические изменения, происходящие в ходе клеточного цикла. При благоприятных для роста условиях общее содержание белка в типичной клетке на протяжении всего цикла увеличивается более или менее непрерывно (рис. 13-7). Синтез РНК тоже происходит с постоянной скоростью, за исключением М-фазы, когда конденсация хромосом, видимо, препятствует транскрипции, так что синтез РНК почти не идет, а образование белка снижается. Анализ синтеза индивидуальных белков (рис. 13-8) показывает, что подавляющее большинство их синтезируется в течение всего цикла. Таким образом, в процессе роста клетки большая часть ее компонентов образуется постепенно и непрерывно - их синтез ненадолго прекращается лишь во время разделения клетки на две.
На фоне этого непрерывного роста происходит ряд резких изменений, связанных с критическими моментами клеточного цикла.
Некоторые из них, такие как начало синтеза ДНК, легко выявляются, тогда как другие обнаружить труднее. Оказалось, например, что для большинства клеток существует критическая точка в фазе G1, когда в их клеточном цикле наступает пауза, если условия среды неблагоприятны для роста. При прохождении этой точки, называемой точкой рестрикции, в клетке происходят внутренние изменения, после которых она должна уже пройти все последующие этапы клеточного цикла в соответствии с жестким временным «расписанием».
Как и следовало ожидать при наличии в цикле ряда критических точек, можно выделить определенные белки (хотя их очень немного), синтез которых резко ускоряется на специфических стадиях цикла (см. рис. 13-8). Например, гистоны, необходимые для построения нового хроматина, синтезируются с высокой скоростью только в S-фазе; это, видимо, относится и к некоторым белкам аппарата репликации ДНК.
Но чем определяется время наступления такого рода критических событий и как они координируются между собой и с непрерывным процессом клеточного роста? Чтобы ответить на эти вопросы, мы вначале посмотрим, что служит пусковым сигналом для синтеза ДНК, т. е. для перехода к S-фазе клеточного цикла.
13.1.4. Синтез ДНК запускается изменением в цитоплазме-появлением активатора S-фазы [4]
Путем добавления надлежащего агента к культуральной среде (разд. 4.3.5) можно вызвать слияние клеток двух синхронизированных клеточных популяций, находящихся в разных фазах клеточного цикла. Результаты такого опыта чрезвычайно информативны.
Когда клетка в S-фазе сливается с клеткой, находящейся на более ранней стадии G1, ядро клетки в фазе G1 немедленно приступает к синтезу ДНК (рис. 13-9, А). Очевидно, что это ядро уже готово
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
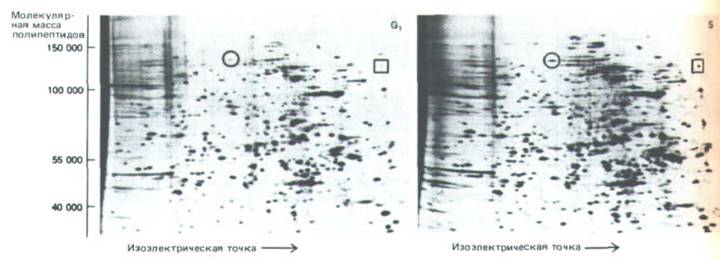
Рис. 13-8. Анализ белков, синтезируемых в фазах G1 и S, с помощью двумерного электрофореза в полиакриламидном геле.
Синхронизированные клетки мышиной лимфомы в течение короткого времени в ранней фазе G 1 или в поздней S-фазе метили смесью радиоактивных аминокислот. На представленных радиоавтографах можно выявить около 1000 вновь синтезированных белков, но только два из них (отмечены кружком и квадратом) образуются в этих двух фазах с существенно разными скоростями. Поскольку гистоны имеют сильный положительный заряд, они мигрируют за пределы геля и здесь не видны. (P. Coffino, V. Е. Groppi, Adv. Cyclic Nucleotide Res. 14: 399-410, 1981.)
к репликации ДНК, но в нормальных клетках G 1 еще отсутствует какой-то сигнал (или ряд сигналов), необходимый для активации механизма синтеза ДНК. По-видимому, в цитоплазме клеток, находящихся в S-фазе, такой сигнальный фактор содержится в большом количестве. Появление такого активатора S-фазы, очевидно, и отмечает границу между фазами G 1 и S в нормальной клетке.
Исчезает ли активатор S-фазы после завершения синтеза ДНК и перехода клетки в фазу G2? Ответ можно опять-таки получить в опытах со слиянием клеток. Когда клетка в фазе G2 сливается с клеткой в фазе G l (рис. 13-9,Б), ядро клетки G 1 не начинает преждевременно синтезировать ДНК; но если такая же клетка G2 сливается с клеткой в S-фазе, то в ядре этой последней репликация ДНК продолжается (рис. 13-9, А). Очевидно, что активатор S-фазы (или какой-то его важный компонент) вскоре после окончания этой фазы исчезает и цитоплазма клеток в фазе G2 уже не содержит ни диффундирующего активатора, ни диффундирующего ингибитора синтеза ДНК.
13.1.5. В каждом цикле весь геном реплицируется только один раз [4, 5]
Как уже говорилось в гл. 9 (разд. 9.3.6), разные части генома реплицируются в разные моменты S-фазы и после этого не могут реплицироваться повторно, так как этому препятствует какое-то химическое

Рис. 13-9. Сводная схема различных результатов слияния двух клеток млекопитающих, находящихся на разных фазах клеточного цикла.
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
изменение, происходящее в каждом участке каждой хромосомы после его репликации. Благодаря такой блокаде повторной репликации при слиянии клетки в фазе G2 с клеткой в S-фазе (рис. 13-9, А) ядро клетки в G2 оказывается нечувствительным к активатору S-фазы и не приступает вновь к синтезу ДНК. Блокада снимается, когда клетки проходят через митоз к началу новой фазы G1.
Однако нужен еще какой-то механизм, который бы гарантировал, что активатор S-фазы будет присутствовать до тех пор, пока не завершится репликация всей ДНК. Как мы видели, опыты со слиянием клеток показывают, что цитоплазматический сигнал, активирующий механизм репликации ДНК в начале S-фазы (активатор S-фазы), исчезает к ее концу. Однако если клетку искусственно блокировать в S-фазе ингибиторами синтеза ДНК, механизм репликации ДНК остается дееспособным и после нормального срока окончания S-фазы, так что в случае удаления ингибитора репликация ДНК возобновляется и доводится до конца. По-видимому, хромосома, не завершившая репликацию, каким-то образом сохраняет механизм репликации в активном состоянии.
Возможно, что за этот эффект ответственны сами репликационные вилки. Как мы видели в гл. 9 (разд. 9.3.1), эти вилки существуют парами: две вилки одной пары движутся в противоположных направлениях от общей начальной точки, и каждая из них прекращает свое существование только тогда, когда она доходит до конца хромосомы или сталкивается с вилкой, движущейся ей навстречу. Таким образом, если хромосома начала репликацию, то будет существовать по крайней мере одна репликационная вилка до тех пор, пока вся хромосома не удвоится полностью. Возможно, что каким-то непонятным образом такая вилка обеспечивает дополнительную выработку активатора S-фазы, эффективно катализирующего образование новых вилок в других участках ДНК. В самом деле, инициация первой пары репликационных вилок могла бы служить пусковым механизмом для начала S-фазы, действующим по принципу «всё или ничего». Такое одиночное событие инициации зависело бы от редкого случайного столкновения между стартовой последовательностью ДНК и молекулой инициатора, присутствующего в низкой концентрации. Действительно, разброс моментов перехода G1→S во времени носит случайный характер, что согласуется с этим предположением (разд. 13.3.3).
ДНК [4, 6]
Ядро, завершившее S-фазу и вступающее в G2, в нормальных условиях конденсирует свои хромосомы и через определенное время после этого вступает в митоз. Однако если синтез ДНК искусственно блокировать, то митоз задержится до тех пор, пока блокада не будет снята и не завершится синтез ДНК. Точно так же после слияния клетки в фазе S с клеткой в фазе G2 ядро последней задерживается на этой стадии, пока другое ядро не «догонит» его, и в конце концов оба ядра вместе вступают в митоз. Проще всего предположить, что задержку митоза вызывает какой-то цитоплазматический сигнал, генерируемый при неполной репликации ДНК. Этот сигнал мог бы быть или не быть идентичным активатору S-фазы;
в любом случае некоторое указание на возможный механизм его появления содержится в том факте, что в случае повреждения клетки в фазе G2
(например, рентгеновским облучением) митоз задерживается до его репарации. Как при репарации, так и при репликации ДНК в клетке должна быть одноцепочечная ДНК;
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
между тем известно, что избыток такой ДНК у бактерий запускает выработку цитоплазматического сигнала, задерживающего клеточное деление (реакция SOS-см. разд. 5.2.9). Возможно, что и в эукариотической клетке одноцепочечная ДНК тоже порождает сигнал задержки М-фазы. Это еще не приближает нас к пониманию природы такого сигнала, если не считать одного очень интересного наблюдения: если в культивируемых клетках млекопитающих искусственно блокировать синтез ДНК (ингибиторами или путем повреждения ДНК), то после добавления в среду кофеина они могут преждевременно приступить к митозу, когда их ДНК еще не реплицировались полностью. На что именно воздействует при этом кофеин, пока не известно.
|
|
|
|
|
Дата добавления: 2015-08-31; Просмотров: 537; Нарушение авторских прав?; Мы поможем в написании вашей работы!