КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Регуляция клеточного деления у многоклеточных организмов
|
|
|
|
У одноклеточных организмов, таких как дрожжи, бактерии или простейшие, отбор благоприятствует тому, чтобы каждая отдельная клетка росла и делилась как можно быстрее. Поэтому скорость деления клеток обычно лимитируется только скоростью поглощения питательных веществ из окружающей среды и переработки их в вещество самой клетки. В отличие от этого у многоклеточного животного клетки специализированы и образуют сложное сообщество, так что главная задача здесь - выживание организма, а не выживание или размножение отдельных его клеток. Для того чтобы многоклеточный организм выжил, некоторые его клетки должны воздержаться от деления, даже если нет недостатка в питательных веществах. Но когда возникает надобность в новых клетках, например при репарации повреждения, ранее не делившиеся клетки должны быстро переключаться на цикл деления; а в случаях непрерывного «износа» ткани скорости новообразования и отмирания клеток всегда должны быть сбалансированы. Поэтому здесь должны существовать сложные регуляторные механизмы более высокого уровня, чем тот, который действует у таких простых организмов, как дрожжи. Этот раздел и посвящен такому «социальному контролю» на уровне отдельной клетки. В гл. 17 и 21 мы познакомимся с тем, как он функционирует в многоклеточной системе для поддержания и обновления тканей тела и какие его нарушения происходят при раке, а в гл. 16 увидим, как еще более сложная система управляет клеточным делением в процессах индивидуального развития.
13.3.1. Различия в частоте деления клеток обусловлены разной длительностью паузы после митоза [16]
Клетки человеческого тела, число которых достигает 1013, делятся с весьма разными скоростями. Нейроны или клетки скелетной мышцы не делятся совсем; другие, например клетки печени, обычно делятся только раз в один или два года, а некоторые эпителиальные клетки кишечника,
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
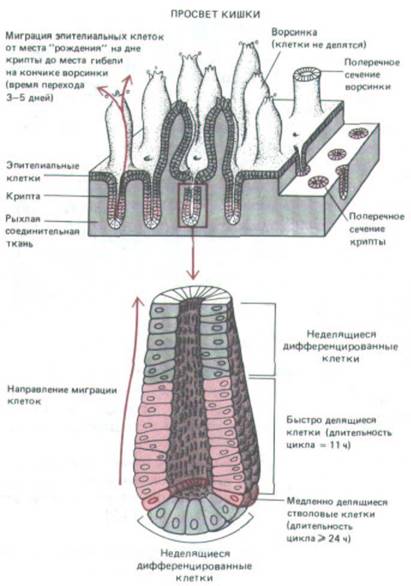
Рис. 13-22. Деление и миграция клеток в эпителиальной выстилке тонкой кишки мыши. Все клеточные деления происходят только в нижней части трубчатых впячиваний эпителия, называемых криптами. Новообразованные клетки перемешаются вверх и образуют эпителий кишечных ворсинок, где они осуществляют переваривание и всасывание питательных веществ из просвета кишки. Большая часть эпителиальных клеток имеет короткий период жизни и слущивается с кончика ворсинки не позднее чем через пять дней после выхода из крипты. Однако кольцо примерно нз 20 медленно делящихся «бессмертных» клеток (их ядра выделены более темным цветом) остаются связанными с основанием крипты.
Эти так называемые стволовые клетки дают при делении две дочерние клетки: в среднем одна из них остается на месте и далее снова функционирует как недифференцированная стволовая клетка, а другая мигрирует наверх, где дифференцируется и входит в состав эпителия ворсинки. (С изменениями из С. S. Pptten, R. Schofield, L G. Lajtha, Biochim. Biophys. Acta 560: 281-299, 1979.)
чтобы обеспечить постоянное обновление внутренней выстилки кишки, делятся чаще чем два раза в сутки (рис. 13-22). Большинство клеток позвоночных располагается где-то в этих временных пределах: они могут делиться, но обычно делают это не так часто. Почти все различия в частоте деления клеток обусловлены разницей в длине промежутка между митозом и S-фазой; медленно делящиеся клетки останавливаются после митоза на недели и даже годы. Наоборот, время, за которое клетка проходит ряд стадий от начала S-фазы до окончания митоза, очень коротко (у млекопитающих обычно от 12 до 24 ч) и удивительно постоянно, каким бы ни был интервал между последовательными делениями.
Время нахождения клеток в непролиферирующем состоянии (так называемой фазе G0) меняется в зависимости не только от их типа, но и от обстоятельств. Половые гормоны побуждают клетки в стенке матки быстро делиться на протяжении нескольких дней в каждом менструальном цикле, чтобы замещать ткань, утраченную при менструации; потеря крови стимулирует пролиферацию предшественников кровяных клеток;
повреждение печени заставляет выжившие клетки этого органа делиться раз или два в сутки, пока не будет возмещена потеря. Точно так же эпителиальные клетки, окружающие рану, приступают к усиленному делению для восстановления поврежденного эпителия (рис. 13-23).
Для регулирования пролиферации клеток каждого типа в соответствии с потребностью существуют тщательно отлаженные и высокоспецифичные механизмы. Однако, хотя важность такой регуляции
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
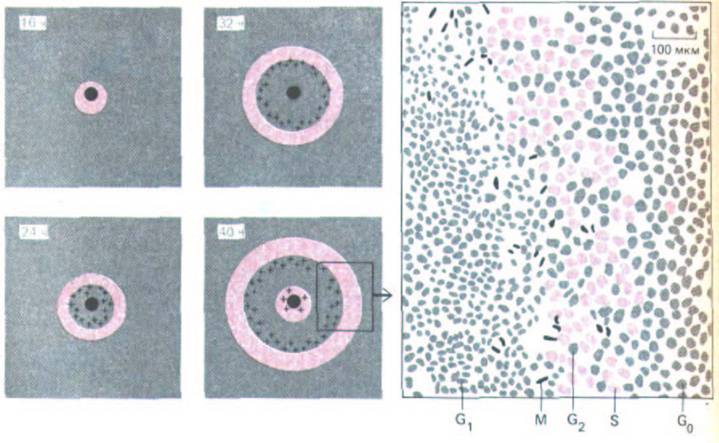
Рис. 13-23. Пролиферация клеток эпителия в ответ на ранение. Эпителий хрусталика повреждали с помощью иглы и спустя определенное время добавляли 3Н-тимидин для мечения клеток в фазе S (выделены цветом); затем вновь фиксировали и приготовляли препараты для р.диоавтографии. На схемах слева участки с клетками в фазе S выделены цветом, а с клетками в фазе М - отмечены крестиками; черное пятно в центре - место нанесения раны. Стимуляция клеточного деления постепенно распространяется от раны, вовлекая в деление покоящиеся клетки в фазе G0, я это приводит к необычно сильной реакции на относительно малое повреждение. На 40-часовом препарате клетки, далеко отстоящие от раны, вступают в фазу S первого цикла деления, тогда как клетки около самой раны вступают в S-фазу второго цикла деления. Рисунок справа соответствует участку, заключенному на схеме слева в прямоугольник; он сделан по фотографии 36-часового препарата, окрашенного для выявления клеточных ядер. (По С. Harding, J. R. Reddan, N.J. Unakar, M. Bagchi, Int. Rev. Cytol. 31: 215-300, 1971.)
очевидна, ее механизмы трудно анализировать в сложном контексте целого организма. Поэтому детальное изучение регуляции клеточного деления обычно проводят на культуре клеток, где легко изменять внешние условия и длительное время наблюдать за клетками.
13.3.2. Когда условия для роста становятся неблагоприятными, клетки животных, так же как и дрожжевые клетки, останавливаются в критической точке в G1 - в точке рестрикции [17]
При изучении клеточного цикла in vitro в большинстве случаев используются стабильные клеточные линии (разд. 4.3.4), способные размножаться неопределенно долго. Это линии, специально отобранные для поддержания в культуре; многие из них - так называемые нетрансформированные клеточные линии - широко используются в качестве моделей пролиферации нормальных соматических клеток.
Фибробласты (такие, как различные типы мышиных клеток ЗТЗ) обычно делятся быстрее, если расположить их в культуральной чашке не слишком плотно и использовать культуральную среду, богатую питательными веществами и содержащую сыворотку - жидкость, получаемую при свертывании крови и очищенную от нерастворимых сгустков и кровяных клеток. При нехватке каких-либо важных питательных веществ, например аминокислот, или при добавлении в среду ингибитора белкового синтеза клетки начинают вести себя примерно так же, как описанные выше дрожжевые клетки при недостатке питания: средняя продолжительность фазы Gt возрастает, но на остальной части клеточного цикла все это почти не сказывается. Как только клетка прошла через G1, она уже неизбежно и без задержки проходит фазы S, G2 и М независимо от условий среды. Эту точку перехода в поздней фазе G1 часто называют точкой рестрикции (R), потому что именно здесь клеточный цикл еще может приостановиться, если внешние условия препятствуют его продолжению. Точка рестрикции соответствует точке старта в клеточном цикле дрожжей; так же как и у дрожжей, она может отчасти служить механизмом, регулирующим размеры клетки. Однако у высших эукариот ее функция более сложна, чем у дрожжей, и в фазе G 1 может быть несколько слегка различающихся точек рестрикции, связанных с различными механизмами контроля клеточной пролиферации.
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
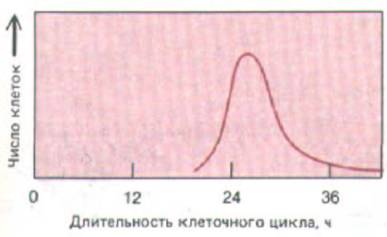
Рис. 13-24. Разброс величин длительности клеточного цикла, наблюдаемый обычно в гомогенной популяции клеток in vitro. Такие данные получают, наблюдая отдельные клетки под микроскопом и прямо отмечая время между последовательными делениями.
13.3.3. Длительность цикла пролиферирующих клеток, по-видимому, имеет вероятностный характер [18]
Индивидуальные клетки, делящиеся в культуре, можно непрерывно наблюдать с помощью цейтраферной киносъемки. Такие наблюдения показывают, что даже у генетически идентичных клеток длительность цикла весьма изменчива (рис. 13-24). Количественный анализ показывает, что время от одного деления до следующего содержит случайно меняющуюся компоненту, причем изменяется она главным образом за счет фазы G1. По-видимому, по мере того как клетки приближаются к точке рестрикции в GJ (рис. 13-25), они должны некоторое время «выждать», прежде чем перейти к оставшейся части цикла, причем для всех клеток вероятность в единицу времени пройти точку R примерно одинакова. Таким образом, клетки ведут себя подобно атомам при радиоактивном распаде; если в первые три часа через точку R прошла половина клеток, в следующие три часа через нее пройдет половина оставшихся клеток, еще через три часа - половина тех, что останутся, и т. д. Возможный механизм, объясняющий такое поведение, был предложен ранее, когда речь шла об образовании активатора S-фазы (разд. 13.1.5). Однако случайные изменения длительности клеточного цикла означают, что первоначально синхронная клеточная популяция через несколько циклов утратит свою синхронность. Это неудобно для исследователей, но может быть выгодно для многоклеточного организма: в противном случае большие клоны клеток могли бы проходить митоз одновременно, а поскольку клетки во время митоза обычно округляются и утрачивают прочную связь друг с другом, это серьезно нарушало бы целостность ткани, состоящей из таких клеток.
|
|
|
|
|
Дата добавления: 2015-08-31; Просмотров: 985; Нарушение авторских прав?; Мы поможем в написании вашей работы!