КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Сбалансированные силы, направленные к противоположным полюсам, удерживают хромосомы в
|
|
|
|
13-28
13-25
13.5.3. Во время митоза хромосомы прикрепляются к мнкротрубочкам своими кинетохорами [37]
Реплицированные хромосомы прикрепляются к митотическому веретену с помощью структур, называемых кинетохорами, В начале М-
фазы каждая хромосома состоит из двух сестринских хроматид, спаренных по всей их длине, но соединенных главным образом возле их центромер- специализированных последовательностей ДНК, необходимых для расхождения хромосом. В поздней профазе на каждой центромере
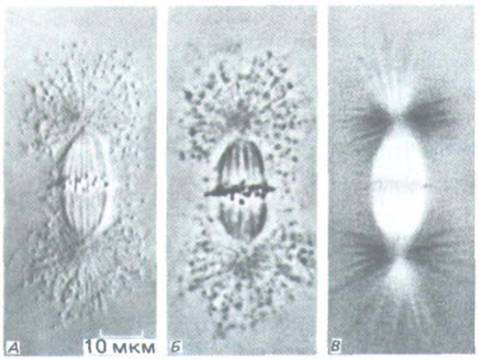
Рис. 13-49. Изолированное метафазное веретено. Использованы три различных метола световой микроскопии: дифференциальный интерференционный контраст (А), фазовый контраст (Б) и микроскопия в поляризованном свете (В). (С любезного разрешения Е. D. Salmon, R.R.
Segall, J. Cell Biol. 86: 355-365, 1980. Repr. by copyright permission of the Rockefeller Univ. Press.)
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
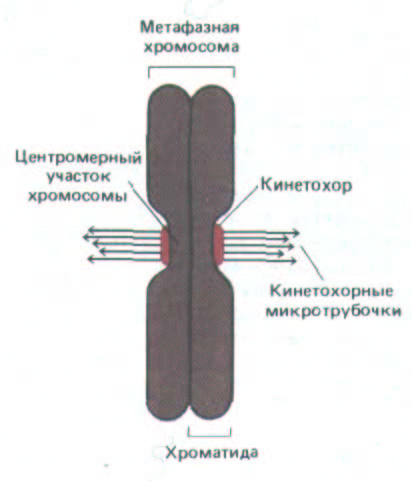
Рис. 13-50. Схематическое изображение метафазной хромосомы с ее двумя сестринскими хроматидамн, к которым прикреплены кинетохорные микротрубочки.
образуется по одному зрелому кинетохору, т. е. имеется теперь два кинетохора (на двух сестринских хроматидах), ориентированных в противоположных направлениях. В метафазе к каждому кинетохору уже прикреплены микротрубочки (рис. 13-50). У большинства организмов кинетохор представляет собой крупный, состоящий из многих белков комплекс, который на электронных микрофотографиях выглядит как пластинчатая трехслойная структура (рис. 13-51). Число микротрубочек, связанных с каждым кинетохором, у разных видов весьма различно:
например, у человека их бывает от 20 до 40, а у дрожжей и некоторых других микроорганизмов -только одна, т.е. одной микротрубочки оказывается достаточно, чтобы тянуть хромосому.
Информация, определяющая специфическую конструкцию кинетохора в специфическом участке хромосомы, должна быть заключена в самой последовательности ДНК центромеры. У дрожжей центромер-ную ДНК можно выявить генетическим методом, благодаря ее способности обеспечивать регулярное наследование плазмид; без ее содействия плазмиды распределяются между дочерними клетками неравномерно и теряются. С помощью методов молекулярной генетики показано, что у дрожжей Saccharomyces ccrevisiae все 17 хромосом содержат разные центромерные последовательности длиной около 110 пар оснований (рис. 13-52); тем не менее во всех этих последовательностях имеются значительные гомологичные участки, которые могут инвертироваться или перемещаться из одной хромосомы в другую без потери функции.
Центромерная последовательность дрожжей связывает специфические белки, которые, видимо, инициируют формирование многобелкового комплекса (кинетохора), а этот комплекс в свою очередь связывается с концом одной микротрубочки. Как полагают, у млекопитающих центромеры состоят из других, гораздо более длинных последовательностей ДНК и формируют более крупные кинегохоры, способные связывать много микротрубочек.
Неожиданная возможность изучать белки кинетохора у млекопитающих появилась, когда стало известно, что у больных некоторыми формами склеродермы (болезни неизвестной природы, связанной с прогрессирующим фиброзом соединительной ткани кожи и других органов), образуются антитела, специфически реагирующие с кинетохорами. Если такие антитела с флуоресцентной меткой использовать для окрашивания делящихся клеток, получается определенный рисунок флуоресцирующих пятен, каждое из которых отмечает положение кинетохора. Такой же пятнистый рисунок создается и в неделящихся клетках; при этом число пятен в клетке соответствует числу ее хромосом (рис. 13-53), и можно думать, что какой-то предшественник кинетохора связан с каждой центромерой даже в интерфазном ядре. Антитела склеродермы сделали также возможным клонирование генов, кодирующих некоторые из
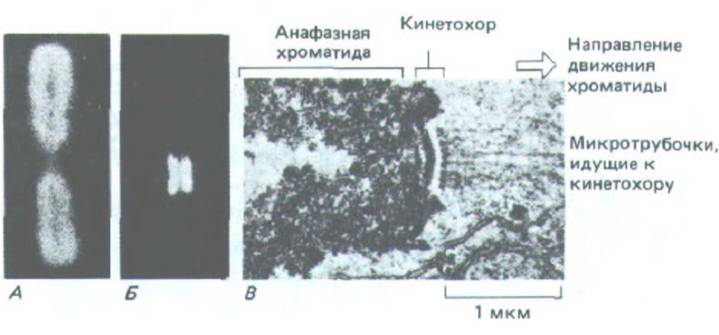
Рис. 13-51. Кинетохоры. В метафазной хромосоме (А), окрашенной аутоантителами человека, реагирующими со специфическими белками кинетохора, выявляются два кинетохора, каждый из которых связан со своей хроматидой (S). На электронной микрофотографии В-
анафазная хроматида с микротрубочками, прикрепленными к кинетохору. Хотя большинство кинетохоров трехслойные, тот, который показан здесь (из зеленой водоросли), имеет необычно сложную структуру с дополнительными слоями. (А и Б с любезного разрешения Bill Brinkley; С - из J.D.
Pickett-Heaps, L. С. Fowke, Aust. J. Biol. Sci. 23: 71-92, 1970. Repr. by permission of CSIRO.)
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.

Рис. 13-52. Последовательность ДНК в типичной центромере дрожжей Saccharomyces cerevisiae. Представленной здесь последовательности достаточно, чтобы обеспечить правильное расхождение хроматид; она служит для сборки белков кинетохора, к которым прикрепляется одна микротрубочка.
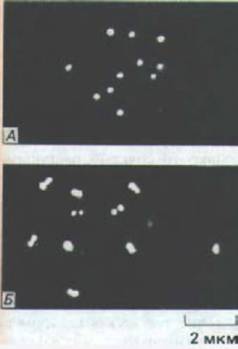
Рис. 13-53. Иммунофлуоресцентное окрашивание кинетохоров в интерфазных клетках с помощью антител, специфически связывающихся с белком кинетохора. В использованных клетках сумчатого относительно мало хромосом. А. В клетках, находящихся в фазе G1, окрашивается один кинетохор на хромосому. Б. В фазе G2 на одну хромосому окрашиваются два кинетохора. (S. L. Brenner, В. В. Brinkley, Cold Spring Harbor Symp. Quant. Biol. 46: 241-254, 1982.)
многочисленных белков, ассоциированных с кинетохорами, так что теперь эти обычно редкие белки можно производить в больших количествах, используя метод рекомбинантной ДНК, а затем изучать их взаимодействие друг с другом, с ДНК и с микротрубочками.
Как микротрубочки и кинетохоры соединяются друг с другом? Их связывание имеет ряд уникальных особенностей. Если химически помеченный тубулин инъецировать в митотическую клетку в метафазе, он будет непрерывно включаться в микротрубочки около точки их прикрепления к кинетохору (рис. 13-54). Как мы увидим позже, в анафазе имеет место обратная реакция: молекулы тубулина отделяются от микротрубочки в участке вблизи кинетохора, так что последний перемещается по направлению к полюсу веретена. Здесь трудно понять то, что кинетохор, несмотря на присоединение и удаление молекул тубулина, сохраняет прочную механическую связь с микротрубочками - ведь именно за эту точку прикрепления они тянут хромосомы сквозь протоплазму. Таким образом, кинетохор, по-видимому, действует наподобие скользящего ошейника, поддерживая боковую связь с субъединицами полимеризованного тубулина около конца микротрубочки и позволяя в то же время добавлять или удалять на этом конце молекулы тубулина (см. ниже рис. 13-61).
13.5.4. По-видимому, кинетохоры захватывают плюс-концы микротрубочек, отходящих от полюса веретена [38]
Распад ядерной оболочки, знаменующий конец профазы и начало лрометафазы, позволяет митотическому веретену взаимодействовать с хромосомами. Конечный результат этого взаимодействия состоит в том, что каждому дочернему ядру будет передано в точности по одной

Рис. 13-54. Опыт, показывающий, что метафазные микротрубочки кинетохора растут с конца, прикрепленного к кинетохору (плюс-
конца). В метафазную клетку млекопитающего in vitro инъецировали тубулин, ковалентне связанный с малой органической молекулой (биотипом).
Спустя 1 мин клетку фиксировали и окрашивали антителами к биотину, связанными с золотыми шариками, а затем приготовляли срезы для электронной микроскопии. Участки микротрубочек, включавшие биотинилированный тубулин в течение минуты после инъекции, усыпаны темными точками золота (цветные стрелки), а участки, существовавшие ранее, не окрашены (черные стрелки). (Фотография любезно предоставлена Luise Evans.)
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
хроматиде каждой хромосомы. В этом процессе распределения хроматид важную роль играют прикрепленные к кинетохорам микротрубочки, которые 1) ориентируют каждую хромосому относительно оси веретена таким образом, чтобы ее два кинетохора были обращены к двум противоположным полюсам клетки, и 2) перемещают каждую хромосому в экваториальную плоскость клетки, где хромосомы образуют метафазную пластинку. В клетках млекопитающих этот процесс занимает 10-20 мин и завершается к концу прометафазы.
Для прометафазы характерна чрезвычайно высокая активность веретена, которое как бы стремится захватить все хромосомы и расположить их в виде метафазной пластинки. И действительно, хромосомы энергично вращаются и движутся туда и сюда между полюсами, так как их кинетохоры присоединены к микротрубочкам, растущим от одного и от другого полюса веретена, и эти микротрубочки тянут их в разные стороны. Первоначальное прикрепление хромосомы обычно происходит тогда, когда она находится у одного из полюсов, и в это время микротрубочки присоединяются только к одному кинетохору; в конце концов и второй кинетохор связывается с микротрубочками, растущими уже от другого полюса. Эти беспорядочные движения хромосом в прометафазе и их окончательная случайная ориентация обеспечивают случайное распределение хроматид между дочерними клетками, что очень важно для перекомбинирования генов во время аналогичного деления ядра в мейозе (разд. 15.2.7).
От полюса отходят только плюс-концы микротрубочек, и именно эти концы связываются с кинетохорами. Таким образом, кинетохор действует как «колпачок» («cap»), в какой-то мере предохраняющий плюс-конец микротрубочки от деполимеризации, точно так же как центромера у полюса веретена предохраняет от деполимеризации' минус-конец. Неудивительно поэтому, что прикрепленные к кинетохору микротрубочки, прикрытые с обоих концов, необычайно стабильны. Другие микротрубочки веретена (называемые полюсными} менее стабильны.
Хотя микротрубочки кинетохора стремятся подтянуть хромосому к соответствующему полюсу (см. ниже), какая-то другая сила, по-
видимому, отталкивает те хромосомы, которые подходят к полюсу слишком близко. Если плечи хромосомы отделить с помощью лазерной микрохирургии от кинетохора, то у них будет заметна тенденция удаляться от ближайшего полюса веретена, даже если они не прикреплены к микротрубочкам или к какой-либо иной клеточной структуре. Одно из возможных объяснений-то, что быстрая полимеризация микротрубочек веретена в направлении от каждого полюса создает «общий поток», который увлекает всякую крупную незакрепленную структуру, такую как плечи хромосом, дальше от полюсов.
13.5.5. Сестринские хроматиды прикрепляются своими кинетохорами к противоположным полюсам веретена [39]
В ранней прометафазе оба кинетохора одной хромосомы могут прикрепиться к нитям от одного и того же полюса веретена. Однако такая или иная аномальная конфигурация, которая привела бы к ошибке в расхождении хромосом, почти всегда исправляется. По-видимому, сбалансированное расположение, при котором сестринские кинетохоры прикреплены к разным полюсам веретена, наиболее стабильно. На возможную причину этого указывают результаты экспериментов, в которых изучался механизм прикрепления хромосом к митотическому веретену.
Изящные опыты, в которых с помощью тончайших стеклянных игл можно было тянуть или толкать хромосомы в живой митотической клетке, показали, что определенный кинетохор не должен быть обяза-
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
тельно направлен к определенному полюсу, подобно концу магнитной стрелки: если хромосому перевернуть, тот же кинетохор может установить связь и с противоположным полюсом. Более того, в прометафазе можно путем микроманипуляции заставить оба кинетохора какой-либо хромосомы связаться с одним и тем же полюсом веретена. Если такая аномальная связь сохранится, то вся хромосома (как пара соединенных сестринских хроматид) будет двигаться к соответствующему полюсу. Как правило, однако, такая связь нестабильна - обычно к хромосоме присоединяются новые микротрубочки от другого полюса с тем, чтобы образовать правильную сбалансированную конфигурацию. С другой стороны, если движению неправильно связанной хромосомы препятствовать стеклянной иглой, то связь этой хромосомы только с одним полюсом становится стабильной: видимо, соединение микротрубочек с кинетохором укрепляется тянущей силой, направленной к полюсу. Поэтому только хромосомы, соединенные с обоими полюсами, будут сохранять связь с микротрубочками и, таким образом, стабильно взаимодействовать с веретеном.
Усилие, развиваемое микротрубочками от разных полюсов, не только стабилизирует взаимодействие этих микротрубочек с кинетохорами, но также в конечном счете приводит каждую хромосому в плоскость метафазной пластинки, о чем сейчас и пойдет речь.
метафазной пластинке [40]
Почему хромосомы в метафазе выстраиваются на равном расстоянии от обоих полюсов веретена, образуя метафазную пластинку?
Опыты по перемещению хромосом стеклянной иглой показывают, что сила, приложенная к кинетохору, пропорциональна длине прикрепленных к нему нитей, т. е. она уменьшается по мере приближения кинетохора к тому полюсу, с которым он соединен (рис. 13-55). Каждая хромосома соединена как бы «пружиной» с каждым из двух полюсов веретена, так что любое смещение к какому-то одному полюсу создает противодействующую силу в обратном направлении. Веретено, образующееся в результате такого взаимодействия в метафазе, показано на рис. 13-
56.
Эти силы продолжают действовать на хромосомы и после того, как те расположились в виде метафазной пластинки. Поэтому и здесь хромосомы совершают колебательные движения в обоих направлениях, поддерживая равновесие этих сил. Если нити, прикрепленные к одному из пары метафазных кинетохоров, разрушить лучем лазера, то вся
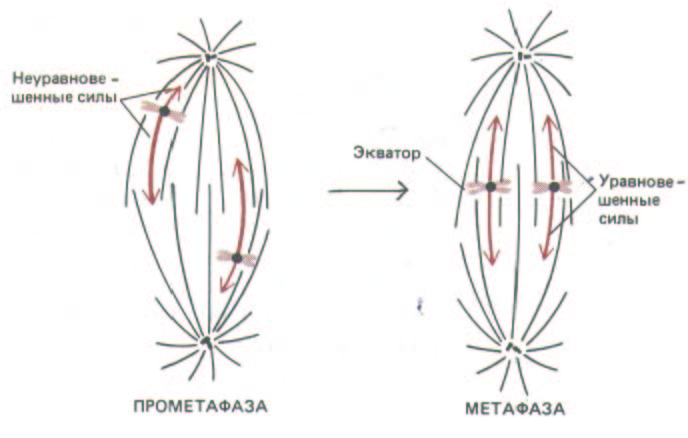
Рис. 13-55. Хромосомы случайным образом попадают в веретено во время прометафазы и в конце концов выстраиваются в экваториальной плоскости веретена, так как сила, действующая на каждый кинетохор, тем меньше, чем он ближе к полюсу. Поэтому хромосомы, оказавшиеся на экваторе, удерживаются там под действием уравновешенных сил притяжения к двум полюсам.
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
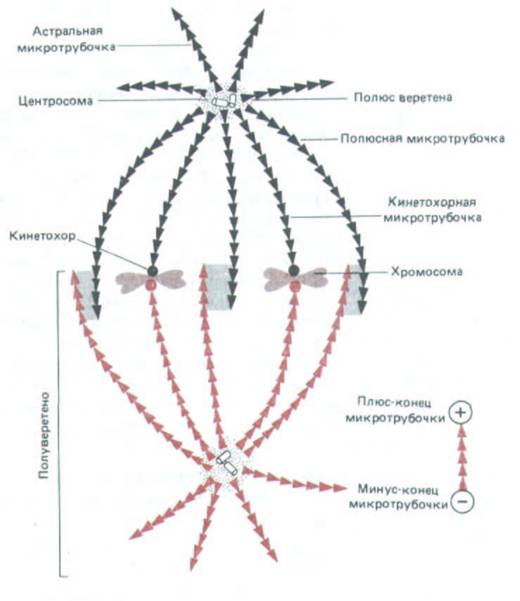
Рис. 13-56. Упрощенная схема мятотического веретена в метафазе. Веретено строится из двух полуверетен (показанных черным и красным цветом), каждое из которых включает кинетохоры, полюсные микротрубочки и микротрубочки звезды. Полярность микротрубочек показана направлением стрелок. Полюсные нити веретена, отходящие от его противоположных полюсов, имеют зону перекрывания (изображена серым цветом), где связанные с микротрубочками белки могут сшивать их. Обратите внимание, что в этой зоне микротрубочки антипараллельны.
хромосома тотчас начнет двигаться к тому полюсу, связь с которым не нарушена. Точно так же, если в метафазе две хроматиды разъединить, то они начнут двигаться к противоположным полюсам, как в анафазе. Судя по этим результатам, как только два кинетохора каждой хромосомы разделятся, хроматиды начинают расходиться к полюсам под действием тех же самых сил, которые раньше привели к образованию метафазной пластинки.
Метафаза занимает значительную часть периода митоза (см. рис. 13-43), как будто клетки выжидают, пока все их хромосомы не расположатся надлежащим образом в экваториальной плоскости. Некоторые эксперименты подкрепляют это представление. Многие клетки останавливаются в митозе на несколько часов или дней, если их обработать такими агентами, как колхицин или винбластин, деполимеризующими микротрубочки; в самом деле, этот способ остановки клеточного цикла широко используют, когда нужно получить большое количество митотических клеток для цитологического анализа их конденсированных хромосом (разд. 9.2.3). После удаления агента митотическое веретено быстро регенерирует, и нередко нормальный митоз возобновляется, как только хромосомы правильно расположатся в метафазной пластинке.
Высказывалось предположение, что хромосома с неприсоединенным кинетохором служит источником диффундирующего фактора, который в норме задерживает переход к анафазе, предоставляя дополнительное время для правильного присоединения. Если такой фактор существует, то при воздействии агентов, разрушающих веретено, следует ожидать появления мощного сигнала, приводящего к продлению метафазы.
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
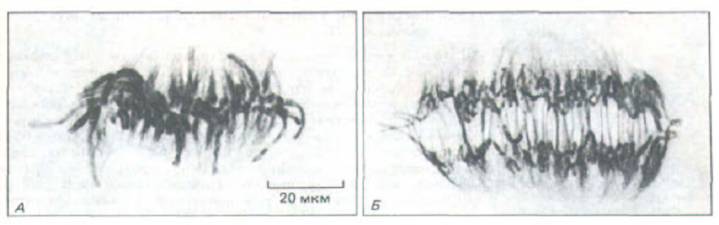
Рис. 13-57. Расхождение хромосом в анафазе клетки из эндосперма Haemanthus. При переходе от метафазы (А) к анафазе (Б) хромосомы растаскиваются полюсными микротрубочками. Клетки окрашены антителами к тубулину, меченными золотом. (С любезного разрешения Andrew Bajer.)
13.5.7. В анафазе сестринские хроматиды внезапно расходятся [41]
Как мы только что видели, метафаза - относительно стабильное состояние, и при обычных условиях многие клетки в течение часа и более пребывают в этой стадии, когда их хромосомы совершают лишь колебательные движения в метафазной пластинке. Анафаза начинается внезапным синхронным расщеплением всех хромосом на сестринские хроматиды, каждая из которых имеет свой кинетохор (рис. 13-57). Сигнал к началу анафазы исходит не от самого веретена, поскольку даже хромосомы, не прикрепленные к веретену, разделяются на хроматиды в то же самое время, что и прикрепленные. Судя по результатам некоторых экспериментов, этот сигнал должен быть связан с повышением концентрации Са2+ в цитозоле. Во-первых, непрерывное наблюдение над клетками, содержащими флуоресцентный индикатор ионов кальция (разд. 4.2.3), показывает, что в некоторых клетках в анафазе происходит быстрое, но кратковременное десятикратное повышение внутриклеточного уровня Са2+. Во-вторых, микроинъекция небольших количеств кальция в культивируемые клетки на стадии метафазы может привести к преждевременному наступлению анафазы. В-третьих, у полюсов веретена обычно видны скопления мембранных пузырьков, и специальная электронно-микроскопическая техника позволяет установить, что эти пузырьки богаты кальцием. Таким образом, возможно, что пузырьки, связанные с веретеном, выделяют С2+ для инициации анафазы (рис. 13-58), подобно тому как саркоплазматический ретикулум высвобождает Са2+ для инициации сокращения скелетной мышцы (разд. 11.1.14).
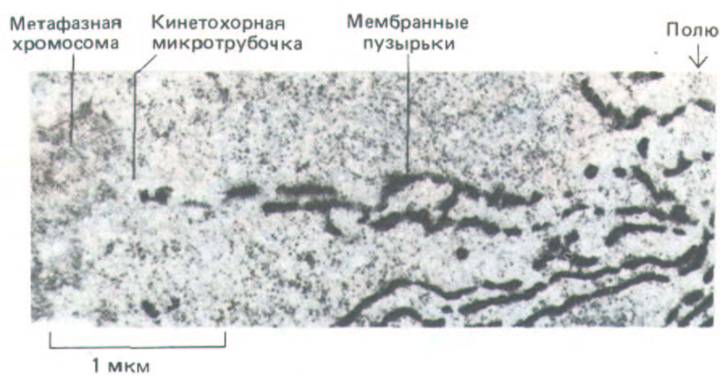
Рис. 13-58. На этой электронной микрофотографии видно скопление специально окрашенных мембранных пузырьков (напоминающих цитоплазматичсский ретикулум) около полюса веретена; пузырьки вытянуты вдоль микротрубочек веретена. (Метафазная клетка из листа ячменя;
фото предоставлено Peter Hepler, из J. Cell Biol. 86: 490-499, 1980, by copyright permission of the Rockefeller Univ. Press.)
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
13.5.8. Расхождение хромосом в анафазе состоит из двух процессов [42]
Как только каждая хромосома расщепилась в ответ на анафазный сигнал, две ее хроматиды начинают двигаться к противоположным полюсам веретена, где они будут включены в ядра новых клеток, По-видимому, это движение - результат двух независимых процессов, происходящих в веретене (рис. 13-59). Первый из них состоит в перемещении хроматид к полюсам и связан с укорочением микротрубочек, прикрепленных к кинетохорам; обычно этот процесс называют анафазой А. Второй процесс - раздвигание самих полюсов, связанное с удлинением полярных микротрубочек и называемое анафазой В. Эти два процесса можно различить по их избирательной чувствительности к некоторым ядам.
Например, низкая концентрация хлоралгидрата предотвращает раздвигание полюсов и удлинение полярных микротрубочек (анафаза В), но не действует ни на микротрубочки кинетохоров, ни на движение хроматид к полюсам (анафаза А). Относительный вклад каждого из этих процессов в окончательное расхождение хромосом существенно различен в зависимости от организма. Например, у клеток млекопитающих анафаза В начинается вскоре после начала движения хроматид к полюсам и заканчивается, когда веретено достигает длины в 1,5-2 раза больше метафазной. У некоторых других клеток, таких как дрожжи, анафаза В начинается только после того, как хроматиды доходят до места своего назначения, а у некоторых простейших анафаза В преобладает и веретено становится в 15 раз длиннее, чем в метафазе.
13.5.9. Во время анафазы А происходит распад микротрубочек, прикрепленных к кинетохорам [43]
При движении хромосом от области метафазной пластинки к полюсам веретена на них воздействуют удивительно большие силы.
Измерения с помощью тонких стеклянных игл дают оценку около 10 -5 дин на
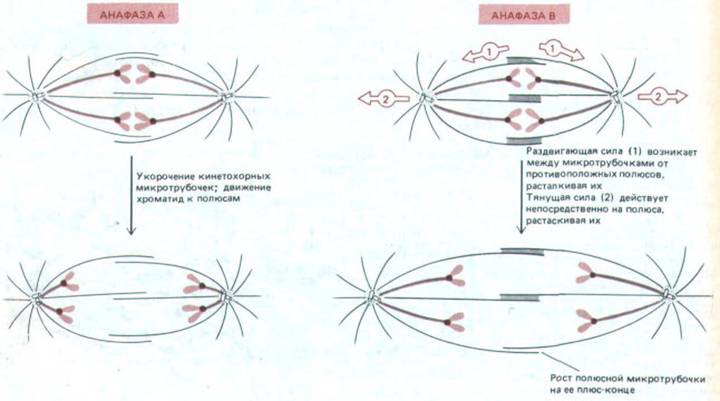
Рис. 13-59. Различные силы, действующие в анафазе при расхождении сестринских хроматид. А. Хроматиды оттягиваются к противоположным полюсам в результате укорочения кинетохорных микротрубочек (движение, называемое анафазой А). Б. В то же время оба полюса веретена отодвигаются дальше друг от друга (движение, называемое анафазой В). Возможно, что силы, обусловливающие анафазу В, подобны тем, которые приводят к расщеплению центросомы и расхождению дочерних центросом с образованием двух полюсов веретена в профазе (см. рис. 13-46). Есть данные о том, что за анафазу В ответственны две отдельные силы: 1} удлинение и скольжение полюсных микротрубочек расталкивают оба полюса, и в то же время 2) другие силы, воздействующие на звезды, тянут полюса в противоположные стороны.
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
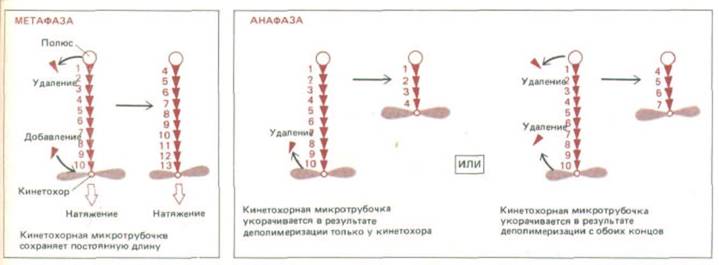
Рис. 13-60. Поведение кинетохорных микротрубочек меняется при переходе от метафазы к анафазе. А. В метафазе на плюс-конце микротрубочки у кинетохора происходит добавление субъединиц тубулина, а на минус-конце у полюса-удаление. Таким образом, субъединицы непрерывно перемещаются в сторону полюса, так что микротрубочки сохраняют постоянную длину и остаются под натяжением. Б. В анафазе натяжение снимается и кинетохор начинает быстро передвигаться по микротрубочке, удаляя при этом субъединицы с ее плюс-конца (слева); в результате этого прикрепленная к нему хроматида перемешается к полюсу веретена. По крайней мере у некоторых организмов движение хроматид частично обусловлено одновременным укорочением микротрубочек также и у полюса (справа).
хромосому, что в 10000 раз больше силы, необходимой для того, чтобы просто продвигать хромосому через цитоплазму с наблюдаемой скоростью.
Очевидно, должен существовать какой-то мощный «мотор» для перемещения хромосом, однако скорость их движения должна лимитироваться не вязкостью среды, а чем-то другим. Как уже отмечалось, тот же «мотор» мог бы осуществлять стягивание хромосом в метафазную пластинку.
По мере того как хромосомы движутся к полюсам, микротрубочки, прикрепленные к их кинетохорам, распадаются, так что в телофазе их почти не видно. Участок, где происходит потеря ими субъединиц, можно определить, введя в клетку меченый тубулин во время метафазы. Было установлено, что меченые субъединицы сначала добавляются к тому концу микротрубочки, который связан с кинетохором, а затем теряются в ходе анафазы А. Это указывает на то, что кинетохор в анафазе как бы «проедает» свой путь к полюсам вдоль своих микротрубочек. В пользу такого вывода говорит и тот факт, что анафазные кинетохоры движутся в сторону стационарной метки, поставленной на микротрубочки. Распад микротрубочек у кинетохоров, полюсов или в обоих этих местах, вероятно, необходим для перемещения хромосом к полюсам (рис. 13-60), так как их движение прекращается, если деполимеризацию микротрубочек блокировать добавлением таксола или D2O.
Механизм, с помощью которого кинетохор, а вместе с ним и хромосома движется по веретену во время анафазы А, остается неизвестным. Две его возможные модели схематически представлены на рис. 13-61. Согласно первой модели, кинегохор при движении вдоль прикрепленной к нему микротрубочки гидролизует АТР, а плюс-конец микротрубочки по мере его обнажения деполимеризуется. В другой модели деполимеризация микротрубочки сама по себе приводит к пассивному движению
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
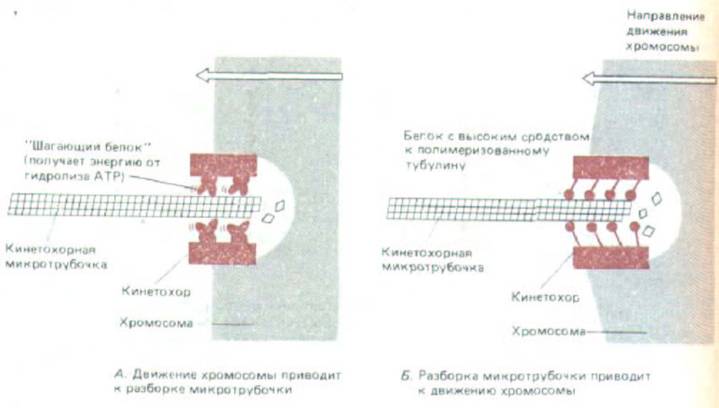
Рис. 13-61. Создание кинетохором силы, движущей хромосому к полюсу в анафазе: две альтернативные модели. А. В кинетохоре имеются «шагающие» белки, сходные с динеином или кинезином; они продвигаются по микротрубочке, используя для этого энергию гидролиза АТР (разд. 10.4.9). Б. Движение хромосом обусловлено распадом микротрубочек: по мере того как субъединицы тубулина диссоциируют, кинетохор, чтобы сохранить связь с микротрубочкой, должен скользить в направлении полюса. Те же механизмы могут использоваться у полюса веретена, который тоже, видимо, способен сохранять связь с микротрубочками, допуская в то же время их контролируемую деполимеризацию (см.
рис. 13-60).
кинетохора. оптимизирующему энергию связывания его с микротрубочкой. Третья возможность, не показанная на рис. 13-61, состоит в том. что микротрубочки не ответственны прямо за возникновение силы, движущей кинетохор к полюсам, а просто регулируют движение, вызываемое какой-то другой структурой. Предполагали, например, что существует система эластичных белковых нитей (возможно, сходных с очень длинными эластичными филаментами поперечнополосатой мышцы - см. разд. 11.1.13), которые связывают кинетохор с полюсом и постепенно подтягивают к нему.
Независимо от природы механизма, создающего силу, нужно еще объяснить драматическое изменение в полимеризации микротрубочек у кинетохора при переходе от метафазы к анафазе (в метафазе преобладает полимеризация, в анафазе - распад, см. рис. 13-60). Возможно, что это связано просто с резким уменьшением тянущего усилия, приложенного к кинетохору, в анафазе; ослабление натяжения могло бы непосредственно изменять динамику полимеризации микротрубочек или же приводить к химическим изменениям в кинетохоре.
13.5.10. В анафазе В, возможно, действуют две различные силы [44]
В анафазе В увеличивается расстояние между двумя полюсами веретена, и в отличие от анафазы А это сопровождается сборкой
микротрубочек. По мере расхождения полюсов полюсные микротрубочки между ними удлиняются, по-видимому путем сборки на своих дистальных плюс-концах.
И удаление полюсов веретена друг от друга в анафазе, и степень перекрывания полюсных микротрубочек в экваториальной зоне сильно варьируют от вида к виду. Зона перекрывания микротрубочек веретена особенно велика у многих диатомовых водорослей (рис. 13-62), у которых митоз происходит внутри ядерной оболочки (разд. 13.5.18), Как показала кропотливая реконструкция трехмерной структуры целых веретен диатомовых водорослей по сотням серийных срезов для электронной микроскопии, у этих водорослей полюсные микротрубочки обоих полуверетен перекрываются в центральной зоне, вблизи от экватора веретена. В анафазе эти две группы антинараллельных микротрубочек, по-
видимому, скользят друг по другу, расходясь в противоположные стороны.
Анафазные движения можно также изучать на лизированных клетках
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.

Рис.. З-62. Эти электронные микрофотографии показывают, как удлиняется веретено и уменьшается степень перекрывания полюсных микротрубочек при митозе у диатомовой водоросли. А. Метафаза. Б. Поздняя анафаза. (С любезного разрешения Jeremy D. Pickett-Heaps.)
диатомей. В такой модельной системе митотическое веретено легко доступно для макромолекул, так что на ней можно испытывать действие различных макромолекулярных агентов, в том числе и специфических антител. Ингибиторы, присоединяющиеся к актину или миозину (в частности, антитела к миозину) не оказывают никакого влияния на движение анафазных хромосом, так что за это движение вряд ли ответственна актомиозиновая система вроде той, которая действует в мышцах. Вместо этого силу здесь могли бы создавать белки, подобные динеину, связанному с микротрубочками в ресничках и жгутиках (разд. 11.3.7), или кинезину, участвующему в быстром аксонном транспорте (разд. 10.4.9). Эти два белка присоединяются к микротрубочкам и вызывают направленное движение за счет гидролиза АТР. но пока не известно, играют ли они существенную роль в митозе.
В клетках высших организмов перед формированием веретена ядерная оболочка разрушается, и поэтому астральные микротрубочки (те, которые направлены от митотического веретена, см. рис. 13-56) могут играть более важную роль в анафазе В, чем у диатомовых. Например, в яйцах некоторых морских беспозвоночных можно разрушить микротрубочки веретена, не блокируя при этом анафазу В. Это позволяет думать, что полюса веретена раздвигаются под влиянием тянущих сил - вероятно, в результате притяжения между астральными микротрубочками и кортексом клетки. Сходные взаимодействия могли бы играть роль и в случаях асимметричного деления клетки (разд. 13.5.13).
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
13.5.11. В телофазе ядерная оболочка образуется сначала вокруг отдельных хромосом [45]
К концу анафазы хромосомы полностью разделяются на две идентичные группы, по одной у каждого полюса веретена. В последней стадии митоза- телофазе -вокруг каждой группы хромосом вновь образуется ядерная оболочка, так что получаются два дочерних интерфазных ядра.
В связи с распадом и восстановлением ядерной оболочки нужно рассмотреть по меньшей мере три ее компонента:
1) наружную и внутреннюю ядерные мембраны, являющиеся продолжением мембран эндоплазматического ретикулума;
2) лежащую под ними ядерную ламину - тонкую двумерную сеть промежуточных филаментов, состоящих из ядерных ламииов, которая взаимодействует с внутренней ядерной мембраной, хроматином и ядерными порами (разд. 11.5.5);
3) ядерные поры, образованные крупными комплексами из недостаточно охарактеризованных белков (разд. 8.3.1).
В профазе многие белки фосфорилируются. Если фосфорилирование молекул гистона Н1, по-видимому, способствует конденсации хромосом (разд. 13.1.10), то фосфорилирование ядерных ламинов участвует в регуляции распада и восстановления ядерной оболочки.
Фосфорилирование ламинов происходит во многих различных участках каждой полипептидной цепи и поэтому приводит к их распаду и, как следствие, к разрушению ядерной ламины. Затем - вероятно, в ответ на другой сигнал - сама ядерная оболочка распадается на, мелкие мембранные пузырьки.
Резкий переход от метафазы к анафазе, по-видимому, приводит к дефосфорилированию многих белков (в том числе молекул гистона Н1
и ламинов), которые были фосфорилированы в профазе. Вскоре после этого, в телофазе, пузырьки ядерной мембраны связываются с поверхностью отдельных хромосом и сливаются, восстанавливая ядерные мембраны, которые лишь частично окружают группы хромосом перед полным восстановлением ядерной оболочки (рис. 13-63); одновременно восстанавливаются и ядерные поры, а дефосфорилированные ламины
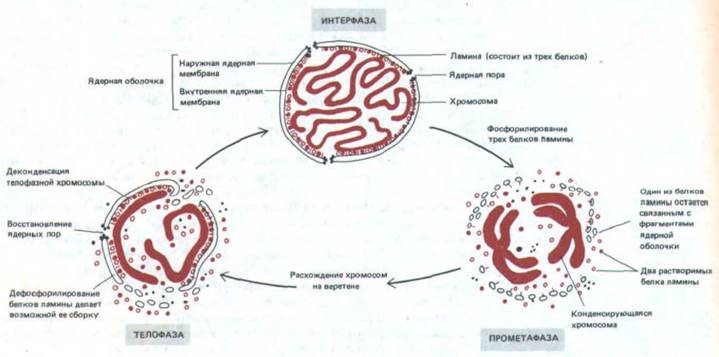
Рис. 13-63. Схема циклических изменений ядерной оболочки во время митоза. В прометафазе ядерные мембраны распадаются на мелкие пузырьки и вновь восстанавливаются в телофазе. Между этими двумя фазами, когда ядерная оболочка разрушена, а ядерные поры и ядерная ламина распались на субъединицы, осуществляются все процессы, в результате которых два набора хромосом расходятся к противоположным полюсам.
Как показано на рисунке, новая ядерная оболочка каждой дочерней клетки образуется в результате слияния мембранных пузырьков вокруг группирующихся индивидуальных хромосом; при этом большая часть цитоплазматических компонентов не попадает в новое ядро.
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
вновь агрегируют, образуя ядерную ламину. Один из белков ламины (ламин В) на протяжении всего митоза остается связанным с фрагментами ядерной мембраны и, возможно, способствует их воссоединению в телофазе. После восстановления ядерной оболочки возобновляется синтез РНК, что ведет к появлению ядрышка (разд. 9.4.19), а хроматин деконденсируется и переходит в дисперсное состояние, характерное для интерфазы.
И распад, и восстановление ядерной структуры могут происходить в неочищенных экстрактах яиц Xenopus, разумеется, если эти экстракты приготовлены из клеток соответствующих стадий клеточного цикла (из мнтотических для распада и из интерфазных для восстановления). В таких экстрактах весь процесс, в котором участвуют ламина, ядерные поры и ядерные мембраны, протекает, по всей видимости, нормально в ответ на циклы фосфорилирования и дефосфорилирования. Таким образом, подобные системы in vitro могут служить тест-объектами при идентификации и очистке белков, катализирующих распад и восстановление ядерной оболочки в клетке, в том числе и белков (таких, как MPF), регулирующих эти процессы. Для восстановления ядра к таким экстрактам нужно добавлять ДНК, причем полное восстановление ядерной оболочки происходит вокруг очищенных молекул ДНК, взятой от любого организма, даже от бактериального вируса. Таким образом, хотя здесь и должны участвовать белки, связывающиеся с ДНК, маловероятно, чтобы при этом распознавались специфические последовательности нуклеотидов.
Интересно, что распад ядерной оболочки не является необходимым для митоза. Действительно, позднее мы увидим, что у низших эукариот ядерная оболочка во время митоза не разрушается; принято говорить, что эти организмы обладают не «открытым», а «закрытым»
веретеном.
13,5.12. Метафазу и интерфазу можно рассматривать как альтернативные «устойчивые» состояния клетки [46]
На рис. 13-64 схематически представлена одна из современных гипотез относительно митотического цикла. В ней принята в некотором смысле точка зрения химика на митоз, где интерфаза и метафаза рассматриваются как два альтернативных «устойчивых» состояния клетки, а другие стадии митоза - просто как необходимые переходные состояния между ними. Предполагается, что в конце интерфазы происходит включение некоего механизма («М-фазного переключателя»), побуждающего клетку проходить через профазу и прометафазу до более устойчивого метафазного состояния. В конце метафазы этот механизм внезапно выключается, и клетка проходит через анафазу и телофазу, возвращаясь к интерфазе, которая при выключенном регуляторе наиболее устойчива.
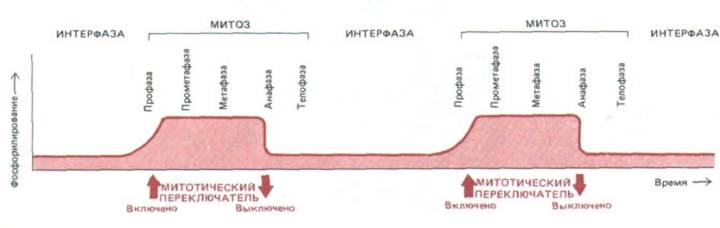
Рис. 13-64. За переход клеток в фазу М и выход из нее, возможно, ответствен некий митотический переключатель с двумя состояниями-
«включено» и «выключено». Согласно этой гипотезе, включение приводит к фосфорилированию многих белков, происходящему только в митотической клетке. Серия структурных изменений, связанных со сборкой веретена, не требует специальных триггеров: скорее это ряд энергетически выгодных этапов на пути к стабильному метафазному состоянию. Это состояние продолжается до тех пор, локa анафазный триггер не поставит переключатель в положение «выключено», а дефосфорилирование белков не восстановит прежние глобальные параметры. Это запускает новую серию структурных изменений (включая расхождение хромосом), что опять приводит клетку к стабильному интерфазному состоянию.
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
Такую точку зрения на митоз подкрепляют данные об изменениях, происходящих с микротрубочками цитоскелета (см. рис. 13-48), а также о внезапных изменениях в активности и степени фосфорилирования некоторых белков, участвующих в митозе, на границах интерфаза/профаза (включение) и метафаза/анафаза (выключение) (см. разд. 13.1.10). Положение «переключателя» могло бы соответствовать уровню активности MPF в клетке (см. обсуждение цикла MPF в разд. 13.1.11 и рис. 13-15).
|
|
|
|
|
Дата добавления: 2015-08-31; Просмотров: 1511; Нарушение авторских прав?; Мы поможем в написании вашей работы!