КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Физиологическая роль и метаболизм серы в растении. Биологический круговорот серы.
|
|
|
|
Сера поглощается растениями только в окисленной форме — в виде сульфат-иона (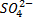 ). Сера содержится в растениях в двух формах — окисленной (сульфат) и восстановленной. Основная часть поглощенного корнями сульфата перемещается в надземную часть растения по сосудам ксилемы к молодым тканям, где он интенсивно включается в обмен веществ.
). Сера содержится в растениях в двух формах — окисленной (сульфат) и восстановленной. Основная часть поглощенного корнями сульфата перемещается в надземную часть растения по сосудам ксилемы к молодым тканям, где он интенсивно включается в обмен веществ.
Попадая в цитоплазму, сульфат восстанавливается с образованием сульфгидрильных групп органических соединений (R–SH). Из листьев сульфат и восстановленные формы серы могут перемещаться как акропетально, так и базипетально в растущие части растения и в запасающие органы. В семенах сера находится преимущественно в органической форме, а в процессе прорастания частично переходит в окисленное состояние. Доля сульфата минимальна в молодых листьях и резко возрастает при их старении в связи с деградацией белков. Сера, как и кальций, не способна к реутилизации и поэтому накапливается в старых тканях растения. Сульфгидрильные группы входят в состав аминокислот, липидов, кофермента А и некоторых других соединений. Сера входит в состав аминокислот цистеина и метионина, которые могут находиться как в свободном виде, так и в составе белков. Потребность в сере особенно высока у растений, богатых белками, например, у бобовых растений и у представителей семейства крестоцветных, в больших количествах синтезирующих серосодержащие горчичные масла, в состав которых входят прототоксины — глюкозинолаты. В клетке наибольшее число тиоловых (–SH) групп приходится на долю трипептидаглутатиона (глу–цис–гли). Одна из основных функций серы связана с формированием третичной структуры белков за счет ковалентных связей дисульфидных мостиков, образуемых между остатками цистеина. Она входит в состав ряда витаминов (липоевой кислоты, биотина, тиамина). Еще одна важная функция серы заключается в поддержании определенного значения окислительно-восстановительного потенциала клетки с помощью обратимых превращений:
2цистеин-SH + ½ O2 -цистеин-S-S-цистеин + H2O
(цистин)
2глутатион-SH +½ O2 -глутатион-S-S-глутатион+ H2O
Недостаточное снабжение растений серой тормозит синтез белков, снижает интенсивность фотосинтеза и скорость ростовых процессов. Внешними симптомами дефицита серы являются бледные и пожелтевшие листья, что проявляется вначале у самых молодых побегов. В почве сера находится в неорганической и органической формах. В большинстве почв преобладают органические формы серы растительных и животных остатков. Основные формы неорганической серы в почве — сернокислые соли кальция, магния и натрия. Они могут находиться в почвенном растворе или быть связанными почвенными коллоидами. В засоленных почвах содержание сульфата натрия может достигать 60% от массы почвы. На затопляемых почвах сера находится в восстановленной форме: (FeS, 2 FeS или 2 HS).
Процесс включения серы в органические соединения состоит из трех этапов. На первом этапе происходит активация сульфата под действием фермента АТФсульфурилазы, который катализирует отщепление пирофосфата от молекулы АТФ и замещение его на сульфат:
Сульфат + АТФ аденозин-5'-фосфосульфат + н Ф-Ф
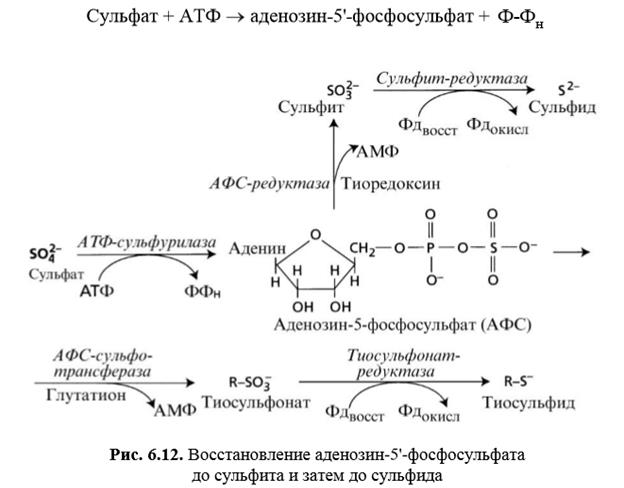
В результате этой реакции относительно инертный сульфат преобразуется в аденозин-5'-фосфосульфат (АФС) и приобретает, таким образом, способность включаться в обмен веществ. На втором этапе ассимиляции происходит восстановление сульфата АФС вначале до сульфита (2 3SO), а затем до сульфида (2 S). В ходе редукции суль- фата АФС может также иметь место образование комплекса ферменттиосульфонат (3 RSO), который далее восстанавливается в тиосульфид (RS) (рис. 6.12). На третьем этапе тиосульфид (RS) или сульфид (2 S) взаимодействуют с О-ацетилсерином с образованием цистеина и уксусной кислоты (рис. 6.13).

Фермент О-ацетилсеринтиолиаза, участвующий в синтезе цистеина, обнаружен в цитоплазме, пластидах и митохондриях растительных клеток. Цистеин в свою очередь служит предшественником еще одной серосодержащей аминокислоты — метионина. В процессе превращения сульфата до цистеина происходит изменение валентности серы от +6 до –4, что требует притока 10 электронов. Донорами электронов на различных этапах восстановления сульфата служат глутатион, ферредоксин (Фд), тиоредоксин, НАД(Ф)H и О-ацетилсерин. Процесс редукции сульфата обычно активнее идет в листьях, поскольку фотохимические реакции, протекающие в хлоропластах, обеспечивают восстановление ферредоксина и тиоредоксина, а в процессе фотодыхания синтезируется серин, из которого образуется О-ацетилсерин, необходимый для синтеза цистеина. Транспорт ассимилированной в листьях серы к местам синтеза белков осуществляется в форме трипептидаглутатиона (глу–цис–гли). Глутатион также является сигнальной молекулой, которая координирует процессы поглощения сульфата корневой системой и его ассимиляции в надземной части растения.
40. Общая роль металлов в жизнедеятельности клетки и целого растения. Металлы макро- и микроэлементы. Металлы и ферменты. Значение калия, кальция, магния, железа, меди, бора и др.
В растительном организме все процессы тесно взаимосвязаны. Исключение из питательной среды какого-либо необходимого элемента быстро вызывает изменение во многих, если не во всех, процессах метаболизма. В связи с этим выделить первичный эффект бывает чрезвычайно трудно. Сказанное относится в первую очередь к тем питательным элементам, которые не входят в состав определенных органических веществ, а играют скорее регуляторную или какую-то иную роль.
В общем виде можно сказать, что питательные элементы имеют следующее значение:
1) входят в состав биологически важных органических веществ;
2) участвуют в создании определенной ионной концентрации, стабилизации макромолекул и коллоидных частиц (электрохимическая роль);
3) участвуют в каталитических реакциях, входя в состав или активируя отдельные ферменты.
Во многих случаях один и тот же элемент может играть разную роль. Некоторые элементы выполняют все три функции.
Макроэлементы.
КАЛИЙ. Он поступает в растение в виде иона К+. Физиологическую роль калия нельзя считать полностью выясненной. Калий не входит ни в одно органическое соединение. Большая часть его (70%) в клетке находится в свободной ионной форме и легко извлекается холодной водой, остальные 30% в адсорбированном состоянии.
• В противоположность кальцию калий снижает вязкость протоплазмы, повышает ее оводненность, увеличивая гидратацию белков. Эта особенность действия калия хорошо проявляется в том, что в его солях плазмолиз имеет выпуклую форму, протоплазма легко отстает от клеточной оболочки. Следовательно, калий является антагонистом кальция.
• Соли калия растворимы и участвуют в регуляции осмотического потенциала клетки. В частности, большое значение имеет К+ в регуляции работы устьиц. Показано, что открытие устьиц на свету связано с накоплением в замыкающих клетках ионов калия. При этом К+ в обмен на Н+ поступает из клеток, окружающих устьичные.
• Корневое давление также во многом зависит от присутствия К+ в пасоке.
• Калий активирует работу многих ферментных систем, например фермент, катализирующий фосфорилирование Сахаров, — гексокиназа, ферменты, катализирующие перенос фосфорной кислоты с пирувата на АДФ (пируваткиназа), а также ферменты, участвующие в образовании АТФ в процессе окислительного фосфорилирования. В связи с этим, по данным Э.И. Выскребенцевой, при недостатке калия резко падает содержание макроэргических фосфатов. Калий активирует и ряд ферментов цикла Кребса. Многие ферменты, участвующие в синтезе белка, требуют для своего действия присутствия калия. Несмотря на необходимость калия для проявления активности ряда ферментов, механизм этой активации до настоящего времени не представляется ясным.
• Недостаток калия замедляет транспорт сахарозы по флоэме. Влияние калия на передвижение органических веществ, по гипотезе Спаннера, проявляется благодаря образованию градиента электрического потенциала на ситовидных пластинках, который возникает при циркуляции калия между ситовидной трубкой и сопровождающими клетками. Влияние К+ на транспорт углеводов определяет его роль в формировании урожая. Под влиянием калия увеличивается накопление крахмала, сахарозы, моносахаридов.
КАЛЬЦИЙ. Поступает в виде иона Са2+. Роль кальция разнообразна.
• Кальций, соединяясь с пектиновыми веществами, дает пектаты кальция, которые являются важнейшей составной частью клеточных оболочек растений. Срединные пластинки, склеивающие клеточные оболочки соседних клеток, состоят по преимуществу из пектатов кальция. При недостатке кальция клеточные оболочки ослизняются, что особенно ярко проявляется в клетках корня. Кальций плохо передвигается по растению, поэтому для предупреждения ослизнения необходимо, чтобы ионы Са2+ непосредственно соприкасались с клетками корня. Сказанное было продемонстрировано в опытах, поставленных по методу изолированных водных культур. В этих опытах одну прядь корней помещали в питательный раствор, содержащий все необходимые питательные вещества; другую прядь корня того же растения — в раствор с исключением кальция.Очень скоро клетки корня, которые находились в растворе без кальция, начали ослизняться и загнивать.
• Кальций повышает вязкость цитоплазмы, что видно на опытах с формами плазмолиза. В солях кальция плазмолиз имеет вогнутую форму, так как более вязкая цитоплазма с трудом отстает от клеточных оболочек.
• Присутствие кальция важно для нормального функционирования мембран. Дефицит кальция приводит к увеличению проницаемости мембран, нарушению их целостности, а соответственно процессов мембранного транспорта.
• Кальций принимает участие в поддержании структуры хромосом, являясь связующим звеном между ДНК и белком. При недостатке кальция наблюдаются повреждения хромосом и нарушение митотического цикла.
• Кальций необходим также для поддержания структуры митохондрий и рибосом, образования ламелл во вновь образующихся клетках.
• Кальций является активатором таких ферментов, как фосфорилаза, аденозинтрифосфатаза, дегидрогеназы, амилазы и др.
• Са2+ служит посредником для реакций растений на внешние и гормональные сигналы, входя в состав сигнальных систем. В этой связи большое значение имеет связывание Са2+ с белком кальмодулином, находящимся в цитозоле. В цитоплазме в обычных условиях поддерживается низкая концентрация кальция. При повышении внутриклеточной концентрации кальция в ответ на сигналы (внутренние и внешние) происходит его связывание с кальмодулином. Кальмодулин регулирует концентрацию Са2+ в клетке по принципу обратной связи. Комплекс Са2+ — кальмодулин способен влиять на активность ферментов, участвующих в синтезе и распаде циклических нуклеотидов (аденилатциклаза, фосфодиэстеразы), Са2+-зависимых протеинкиназ, Са2+-АТФазы. Кальмодулин влияет на активность структурных белков цитоскелета и таким образом контролирует перемещение органелл внутри клетки, изменение формы клетки, образование веретена деления.
• Кальций участвует в образовании клеточной стенки и росте растяжением.
• Кальций реагирует с различными органическими кислотами, давая соли, и тем самым является в определенной мере регулятором значения рН клеточного сока. Нейтрализуя щавелевую кислоту, образует характерные кристаллы щавелевокислого кальция.
МАГНИЙ. Магний поступает в растение в виде иона Mg2+.
• Магний входит в состав основного пигмента зеленых листьев — хлорофилла.
• Магний поддерживает структуру рибосом, связывая РНК и белок. Большая и малая субъединицы рибосом ассоциируют вместе лишь в присутствии магния, который также необходим для формирования полисом и активации аминокислот. Поэтому синтез белка не идет при недостатке магния, а тем более в его отсутствие.
• Магний является активатором многих ферментов. Важной особенностью магния является то, что он связывает фермент с субстратом по типу хелатной связи (клешневидная связь между органическим веществом и катионом). Так, например, присоединяясь к пирофосфатной группе, магний связывает АТФ с соответствующими ферментами. В связи с этим все реакции, включающие перенос фосфатной группы (большинство реакций синтеза, а также многие реакции энергетического обмена), требуют присутствия магния. Магний активирует такие ферменты, как ДНК- и РНК-полимеразы, аденозинтрифосфатазу, глютаматсинтетазу; ферменты, катализирующие перенос карбоксильной группы,— реакции карбоксилирования и декарбоксилирования; ферменты гликолиза и цикла Кребса, молочнокислого и спиртового брожений. В ряде случаев влияние магния на работу ферментов определяется тем, что он реагирует с продуктами реакции, сдвигая равновесие в сторону их образования.
• Магний может также инактивировать ряд ингибиторов ферментативных реакций.
ЖЕЛЕЗО. Несмотря на ничтожное содержание, его роль рассматривается вместе с макроэлементами. Железо поступает в растение в виде Fe3+, а транспортируется в листья по ксилеме в виде цитрата железа (III).
Роль железа в большинстве случаев связана с его способностью переходить из окисленной формы (Fe3+) в восстановленную (Fe2+) и обратно.
• Железо входит в состав каталитических центров многих окислительно-восстановительных ферментов. В виде геминовой группировки оно входит в состав таких ферментов, какцитохромы, цитохромоксидаза, нитратредуктаза, нитритредуктаза, леггемоглобин, каталаза и пероксидаза. Цитохромная система является необходимым компонентом дыхательной и фотосинтетической электронтранспортной цепи. В силу этого при недостатке железа тормозятся оба этих важнейших процесса. Кроме того, целый ряд ферментов содержит железо в негемовой форме. К таким ферментам относятся некоторые флавопротеиды, нитрогеназа, железосодержащий белок ферредоксин, фитоферритин и др. Фитоферритин — является металлопротеидом, в виде которого железо аккумулируется в клетке.
• Железо необходимо для образования хлорофилла. При этом железо катализирует образование предшественников хлорофилла 5-аминолевулиновой кислоты и протопорфиринов. Предполагают, что железо играет роль в образовании белков хлоропластов. При недостатке железа нет условий для образования таких важнейших компонентов хлоропластов, как цитохромы, ферредоксин и некоторые другие. Возможно, это косвенно влияет на образование хлорофилла.
• В хлоропластах железо в негемовой форме входит в состав реакционных центров фотосистем I и II.
Микроэлементы
МЕДЬ. Поступает в растение в виде иона Сu2+ или Сu+.
• Медь входит непосредственно в состав ряда ферментных систем, относящихся к группе оксидаз, таких, как полифенолоксидаза, аскорбатоксидаза, цитохромоксидаза. В этих ферментах медь соединена с белком, по-видимому, через SH-группы. Полифенолоксидаза и аскорбатоксидаза осуществляют окисление фенолов и аскорбиновой кислоты, а цитохромоксидаза входит в состав дыхательной цепи митохондрий.
• Ряд ферментов медь активирует, в частности нитратредуктазу, а также протеазы. Это объясняет роль меди в азотном обмене.
• Большая часть меди (75% от всего содержания меди в листьях) концентрируется в хлоропластах. В хлоропластах сосредоточен и медьсодержащий белок синего цвета — пластоцианин. Содержание меди в пластоцианине составляет 0,57%. Медь, подобно железу и марганцу, обладает способностью к обратимому окислению и восстановлению: Сu2+ + е —>Сu+. Именно поэтому пластоцианин участвует в переносе электронов от ФСII к ФСI. При дефиците меди снижается активность первой фотосистемы. В связи с этим понятно значение меди для процесса фотосинтеза.
МАРГАНЕЦ. Поступает в растение в виде ионов Мп2+. В растении марганец находится в разной степени окисления (Мп2+, Мп3+, Мп4+).
Марганец характеризуется высоким показателем окислительно-восстановительного потенциала. С этим связано значение этого элемента в реакциях биологического окисления.
• Он необходим для нормального протекания фотосинтеза, поскольку входит в состав активного центра кислородовыделяющего комплекса фотосистемы II и осуществляет разложение воды и выделение кислорода: 2Мп4+ + 2Н20 -> 2Мп2+ + 4Н+ + 02.
• Кроме того, марганец участвует в восстановлении С02, играет роль в поддержании структуры хлоропластов. В отсутствие марганца хлорофилл быстро разрушается на свету.
• Марганец активирует более 35 ферментов, участвующих в реакциях окисления-восстановления, декарбоксилирования и гидролиза. В том числе ферменты, катализирующие реакции цикла Кребса (дегидрогеназа яблочной кислоты, лимонной кислоты, декарбоксилаза щавелевоуксусной кислоты и др.). В связи с этим понятно большое значение марганца для процесса дыхания, особенно его аэробной фазы.
• Марганец участвует в азотном обмене в восстановлении нитратов до аммиака, поскольку в этом процессе участвуют ферменты, некоторые из которых зависимы от марганца. В связи с этим у растений, испытывающих недостаток марганца, затруднено использование нитратов в качестве источника азотного питания.
• Марганец связан с синтезом белка через регуляцию активности ДНК-полимеразы и РНК-полимеразы.
• Марганец активирует ферменты, участвующие в окислении важнейшего фитогормона — ауксина.
ЦИНК. Поступает в растение в виде ионов Zn2+.
В растениях цинк не участвует в окислительно-восстановительных реакциях, поскольку не меняет степень окисления.
• Он входит в состав более 30 ферментов, в т. ч. фосфатазы, карбоангидразы, алкогольдегидрогеназа, РНК-полимераза и др. Карбоангидраза катализирует разложение гидрата окиси углерода на воду и углекислый газ. Эта реакция важна для процесса фотосинтеза. Углекислый газ, поступая в клетку, растворяется в воде, образуя Н2С03: С02 + Н20 <->НС03 + Н+. Фермент карбоангидраза, катализируя высвобождение С02 из гидрата окиси углерода, способствует его использованию в процессе фотосинтеза. Кроме того, цинк активирует такие ферменты, как енолаза, альдолаза, гексокиназа, триозофосфатдегидрогеназа. В этой связи понятно значение цинка для процессов дыхания и фотосинтеза.
• Цинк играет важную роль при образовании фитогормона ауксина. Это связано с тем, что цинк, повышая активность триптофансинтетазы, влияет на образование аминокислоты триптофана — предшественника ауксина. Внесение цинка повышает содержание ауксинов и заметно сказывается на темпах роста растений.
• При дефиците цинка возрастает проницаемость мембран, что свидетельствует о роли этого элемента в структуре мембран, в поддержании их интеграции.
• Цинк влияет на белковый синтез, на активность РНКазы. Обнаружены белки, содержащие цинк и участвующие в репликации ДНК и транскрипции.
• Цинк входит в состав одного из факторов регуляции транскрипции в соединении с остатками гистидина и цистеина («цинковые пальцы»).
МОЛИБДЕН. Поступает в растения в виде аниона Мо042-.
• Молибден входит в состав более 20 ферментов, выполняя при этом не только каталитическую, но и структурную функцию.
• Молибден вместе с железом входит в состав активного центра ферментного комплекса нитрогеназы в виде Mo-Fe-белок и участвует в фиксации азота атмосферы различными микроорганизмами. При недостатке молибдена происходят заметные изменения в азотном обмене растений — наблюдается уменьшение синтеза белка при одновременном падении содержания аминокислот и амидов. Нарушения в азотном обмене особенно проявляются на фоне питания растений нитратами. Это связано с тем, что молибден входит в активный центр фермента, восстанавливающего нитраты до нитритов,— нитратредуктазу. Нитратредуктаза — это флавопротеид, простетической группой которого является флавинадениндинуклеотид (ФАД). При восстановлении нитратов молибден действует как переносчик электронов от ФАД к нитрату, при этом N03- переходит в N02-, а Мо5+ — в Мо6+. Образование нитратредуктазы является одним из немногих примеров адаптивного синтеза ферментов в растительном организме. Этот фермент образуется, когда в среде имеются нитраты и молибден. Активность нитратредуктазы возрастает в 10 раз и более при питании растений нитратами по сравнению с аммиаком, причем появление фермента происходит уже через 1—3 ч после внесения в среду нитратов.По-видимому, молибден обладает и другими функциями, так как необходим растению и в условиях достаточного уровня аммиачного питания.
• При недостатке молибдена резко падает содержание аскорбиновой кислоты.
• При отсутствии молибдена наблюдаются нарушения в фосфорном обмене растений.
• Со способностью молибдена к комплексообразованию связано влияние этого элемента на стабилизацию структуры нуклеиновых кислот.
БОР. Поступает в растение в виде аниона борной кислоты — В033-. Роль бора выяснена далеко не достаточно. Это связано с тем, что бор, в отличие от большинства других микроэлементов, не входит в состав ни одного фермента и не является активатором ферментов.
• Большое значение для осуществления функции бора имеет его способность давать комплексные соединения. Комплексы с борной кислотой образуют простые сахара, полисахариды, спирты, фенольные соединения и др. В этой связи можно предположить, что бор влияет на скорость ферментативных реакций через субстраты, на которые действуют ферменты.
• Комплексы органических соединений с борной кислотой могут иметь и иное значение. Так, способность бора образовывать комплексы с углеводами оказывает влияние на клеточную оболочку, регулируя ориентацию мицелл целлюлозы, что способствует ее большей эластичности (Е.В. Бобко). У растений, испытывающих недостаток бора, наблюдается быстрая потеря эластичности клеточных оболочек, что, в свою очередь, связано с более жесткой ориентацией мицелл целлюлозы. Бор играет роль в поддержании структуры мембран.
• При недостатке бора подавляется активность Н+-помпы, повышается проницаемость мембран.Комплексы Сахаров с бором — сахаробораты, по-видимому, легче проникают через мембраны и быстрее передвигаются по растению. При недостатке бора сахара накапливаются в листьях, и их отток резко тормозится. Определения, проведенные с нанесением меченой сахарозы на листья, показали, что при небольшом добавлении борной кислоты отток сахарозы идет значительно быстрее. Сходные результаты были получены в опытах, в которых лист экспонировался в атмосфере, содержащей меченый С02. Образовавшиеся при этом меченые продукты фотосинтеза оттекали значительно быстрее у растений, получивших бор.
• Показано (М.Я. Школьник), что при недостатке бора нарушается синтез нуклеиновых кислот. В боронедостаточных растениях заторможен процесс аминирования органических кислот.
• Бор может выступать как ингибитор активности ряда ферментов, в первую очередь катализирующих образование фенольных соединений.
• При недостатке бора наблюдается накопление кофейной и хлорогеновой кислот, которые считаются ингибиторами роста растений.
• Бор усиливает рост пыльцевых трубок, прорастание пыльцы. В этой связи опрыскивание бором способствует оплодотворению.
КОБАЛЬТ. Находится в тканях растений в ионной (Со2+, Со3+) и комплексной форме.
• Особенно кобальт необходим бобовым растениям, поскольку участвует в фиксации атмосферного азота. Кобальт входит в состав кобаламина (витамин В12 и его производные), который синтезируется бактериями в клубеньках бобовых растений, а также в состав ферментов у азотфиксирующих организмов, участвующих в синтезе метионина, ДНК и делении клеток бактерий.
• При дефиците кобальта подавляется синтез леггемоглобина, снижается синтез белка, и уменьшаются размеры бактероидов. Это говорит в пользу необходимости кобальта. Установлена потребность в кобальте для высших растений, не способных к азотфиксации.
• Показано влияние кобальта на функционирование фотосинтетического аппарата, синтез белка, его связь с ауксиновым обменом. Трудность решения вопроса о необходимости кобальта для всех растений заключается в том, что потребность в нем чрезвычайно мала.
НИКЕЛЬ. Поступает в растения в виде иона Ni2+, но может также находиться в виде Ni+ и Ni3+, Роль никеля для высших растений как микроэлемента была доказана недавно. До этого считали никель необходимым микроэлементом многих бактерий.
• У высших растений никель входит в состав фермента уреазы, который осуществляет реакцию разложения мочевины. Показано, что в растениях, обеспеченных никелем, активность уреазы выше и соответственно ниже содержание мочевины по сравнению с необеспеченными.
• Никель активирует ряд ферментов, в т. ч. нитратредуктазу и другие, оказывает стабилизирующее влияние на структуру рибосом.
• Имеются еще и такие элементы, которые усиливают рост лишь определенных групп растений.
НАТРИЙ. Для роста некоторых растений засоленных почв (галофитов) оказывается полезным натрий. Необходимость натрия проявляется у растений С4 и САМ. У этих растений показана необходимость натрия для регенерации ФЕП при карбоксилировании. Недостаток натрия у этих растений приводит к хлорозу и некрозам, а также тормозит развитие цветка. В натрии нуждаются и многие С3-растения. Показано, что этот элемент улучшает рост растяжением и выполняет осморегулирующую функцию, подобно калию. Благоприятное влияние оказывает натрий на рост сахарной свеклы и цианобактерий.
|
|
|
|
|
Дата добавления: 2017-02-01; Просмотров: 173; Нарушение авторских прав?; Мы поможем в написании вашей работы!