КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
В зависимости от условий среды
|
|
|
|
Изменение активности ферментов
На каталитические свойства ферментных белков оказывают влияние многие химические и физические факторы, способные воздействовать на структуру активного центра, скорость образования и распада фермент- субстратного комплекса, состояние ионизации молекул субстрата и определенных группировок фермента: температура, рН и ионный состав среды, присутствие активаторов и ингибиторов, окислительно-восстановительный потенциал среды, концентрация субстрата и фермента, влияние внешнего электрического поля и др. Действие этих факторов оказывается возможным благодаря тому, что ферменты являются белками, которые довольно легко перестраивают пространственную структуру молекулы под влиянием условий внешней среды и, следовательно, проявляют высокую лабильность в изменении своих физико-химических свойств. При оценке влияния факторов внешней среды на каталитические свойства ферментов следует также учитывать, что в каждом конкретном случае определяется активность того набора изоферментов, которые определяют специфичность обмена веществ в исследуемом биологическом объекте.
Влияние температуры. Ферментативные реакции могут происходить в интервале температур от 0оС (точка замерзания воды) до 70-80оС (тепловая денатурация белков высших организмов). При повышении температуры увеличиваются скорости химических реакций, в том числе и скорость образования фермент–субстратных комплексов, поэтому активность ферментов возрастает, вследствие чего происходит интенсификация процессов жизнедеятельности растений.
Ускорение реакций под воздействием повышенной температуры обу-словлено тем, что снижается энергия активации как субстрата, так и фер-мента. В интервале температур 0-40оС повышение температуры на 10оС увеличивает скорость ферментативных реакций в 1,4 – 2 раза. Наиболее высокая активность ферментов наблюдается при температурах 40-50оС. При температурах свыше 50оС активность ферментов понижается, так как начинается процесс тепловой денатурации ферментных белков, вызывающий необратимое разрушение четвертичной, третичной и вторичной структур белковых молекул. Полная денатурация растительных белков в растворе, как указывалось ранее, происходит при температурах 70-80оС.
Однако в негидратированном (сухом) состоянии белки способны сохранять нативные свойства и при более высокой температуре, что используется в технологиях быстрого высушивания зерна и семян, без снижения их всхожести. Не происходит необратимой инактивации ферментов также и при замораживании растительных тканей. При понижении температуры ниже точки замерзания физиологической среды прекращается действие ферментов, однако при повышении температуры их каталитическая активность восстанавливается.
Влияние рН среды. Каждая молекулярная форма фермента (изофермент) проявляет каталитическую активность в узком интервале рН среды, в связи с тем, что от концентрации водородных ионов зависит состояние ионизации молекулы субстрата и активных группировок в активном центре ферментного белка, которое обеспечивает каталитическое действие фермента. Кроме того, концентрация ионов водорода влияет на конформацию активного центра. Поэтому даже небольшой сдвиг реакции среды от оптимального значения изменяет заряд кислотных и основных групп фермента и субстрата и конформацию активного центра фермента, вследствие чего резко понижается каталитическая активность ферментной молекулы. Однако ферменты в клетках растений представлены наборами изоферментов, каждый из которых проявляет каталитическую активность при разных значениях рН среды и в результате действия всего набора изоферментов обеспечивается ход катализируемой реакции в широком диапазоне изменения рН среды. Зависимость активности фермента от рН среды показана на рис. 22.
Влияние концентрации фермента и субстрата. Если в физиологической среде содержится много субстрата и мало ферментного белка, то скорость превращения субстрата в продукты реакции будет низкой, так как каждая молекула фермента способна катализировать превращение определенного количества субстрата в единицу времени. При увеличении числа ферментных молекул (т.е. концентрации фермента) в среде скорость ферментативной реакции будет возрастать до тех пор, пока достаточно субстрата для полной реализации молярной активности фермента. При дальнейшем увеличении концентрации фермента скорость реакции уже не возрастает. Следовательно, скорость ферментативной реакции пропорционально зависит от количества фермента в среде только при высокой концентрации субстрата.
Скорость реакции, катализируемой ферментом, также зависит от концентрации субстрата в физиологической среде при условии, когда количество фермента остается неизменным. Если в среде находится определенное количество фермента и в нее добавляется субстрат, то вначале скорость ферментативной реакции будет возрастать пропорционально увеличению концентрации субстрата. Однако в дальнейшем по мере увеличения концентрации субстрата темпы нарастания скорости реакции замедляются и при определенном количестве субстрата в среде скорость реакции достигает максимального уровня, при котором полностью реализуется молярная активность фермента (рис. 23). С этого момента скорость реакции уже не будет возрастать при увеличении количества субстрата в среде, так как лимитирующим фактором, определяющим дальнейший ход реакции, уже становится концентрация фермента.
Зависимость скорости ферментативной реакции (V) от концентрации субстрата может быть выражена уравнением:
[ S]
V = Vmax ×¾¾¾¾
Km +[S ]
В этом уравнении [S] - концентрация субстрата; Vmax – максимальная скорость реакции при высокой концентрации субстрата, когда в каждый момент времени все молекулы фермента находятся в соединении с субстратом (насыщение фермента субстратом). Кm – константа Михаэлиса, названная именем Л. Михаэлиса, который внес большой вклад в разработку теории ферментативной кинетики.
Константа Михаэлиса выражает сродство фермента к субстрату и является важной характеристикой ферментного белка. Чем меньше константа Михаэлиса, тем выше молярная активность фермента и тем интенсивней происходит ферментативный катализ. Из приведенного выше уравнения можно определить значение константы Михаэлиса:
Vmax Vmax
Km = [S] (¾¾ – 1); при V = ¾¾ Km = [S]
V 2
Следовательно константа Михаэлиса численно равна концентрации субстрата, при которой скорость реакции, катализируемой ферментом, достигает половину от максимальной.
Для определения константы Михаэлиса экспериментальным путем с использованием чистого фермента находят скорость катализируемой реакции при разных значениях концентрации субстрата, включая максимальную скорость реакции. Затем изменение скорости ферментативной реакции в зависимости от концентрации субстрата выражают в виде графика, как показано на рис. 23. Если на этом графике отметить скорость, равную половине от максимальной, и опустить перпендикуляр на ось абсцисс, то он укажет концентрацию субстрата, численно равную константе Михаэлиса.
Активаторы ферментов. Для поддержания молекулы фермента в активном состоянии необходимо наличие в среде определенных ионов и некоторых других соединений, называемых активаторами ферментов. Роль активаторов заключается в том, что они способны переводить в активное состояние определенные группировки в каталитическом центре молекулы фермента и таким образом участвовать в каталитическом действии ферментного белка. Так, например, протеолитические ферменты, катализирующие гидролитическое расщепление белков, активируются HCN, H2S, а также веществами, содержащими сульфгидрильные группы (восстановленный глютатион, цистеин).
Многие активаторы выполняют роль дополнительного субстрата, который взаимодействуя с истинным субстратом, переводит его в активное состояние и тем самым ускоряет образование фермент-субстратного комплекса. Примером такого взаимодействия могут служить ферментативные реакции с участием АТФ и ионов магния, в которых основной субстрат АТФ переводится в активное состояние путем образования комплекса с катионом магния (Мg АТФ2–, см. стр. 199).
На каталитическую активность ферментов оказывает влияние ионный состав среды, способствующий формированию молекулами фермента и субстрата специфической пространственной структуры, которая позволяет этим молекулам активно взаимодействовать, в результате увеличивается скорость образования фермент-субстратного комплекса и в целом происходит ускорение ферментативной реакции.
Многие ферменты активируются в присутствии ионов металлов. Так, катионы магния (Mg2+) являются активаторами фермента фосфопируватгидратазы, катализирующей отщепление молекулы воды от 2-фосфогли-цериновой кислоты в процессе дыхания, а также ферментов, катализирующих реакции фосфорилирования, в которых происходит перенос остатков ортофосфорной кислоты от АТФ на различные субстраты. Катионы кальция (Са2+) активируют растительные липазы, катализирующие гидролиз ацилглицеринов, и a-амилазы, ускоряющие распад крахмала. Катионы калия (К+) активируют многие ферменты углеводного обмена. Фермент аргиназа, катализирующий гидролитическое расщепление аргинина на цитруллин и мочевину, повышает свою активность в присутствии катионов Со2+, Мn2+, Ni2+.
Известны также ферменты, активность которых повышается в присутствии неорганических анионов: Cl‾, Br‾, I‾, H2PO4‾, НСО3‾ и др. Так, активаторами амилаз, катализирующих гидролиз крахмала, являются ионы галогенов.
При изучении влияния активаторов на активность ферментов было выяснено, что максимальная каталитическая активность ферментных молекул наблюдается при определенной концентрации активатора, при ее уменьшении или увеличении по сравнению с оптимальным уровнем каталитические свойства фермента ослабляются.
Таким образом, для проявления максимальной активности ферментов необходимо наличие в физиологической среде, в которой функционирует фермент, определенного набора специфических активаторов, содержащихся в оптимальной концентрации.
Ингибиторы ферментов. Это вещества, которые подавляют действие ферментов. Под влиянием ингибитора скорость ферментативной реакции замедляется или ферментативное превращение полностью прекращается. Процесс воздействия ингибитора на ферментные белки называют ингибированием ферментов. Различают обратимое и необратимое ингибирование и соответственно ингибиторы обратимого и необратимого действия.
При обратимом ингибировании не происходит безвозвратной потери каталитической активности фермента, так как ингибитор не разрушает пространственной структуры ферментного белка и после отделения ингибитора от фермента активность последнего восстанавливается. Различают два вида ингибиторов, вызывающих обратимое ингибирование, - конкурентные и неконкурентные ингибиторы.
Конкурентные ингибиторы – вещества структурно родственные субстрату, вследствие чего они, как и молекулы субстрата, способны связываться с одним из участков в активном центре фермента, но при этом не могут подвергаться превращению под действием фермента. Молекулы фермента, связанные с ингибитором, не могут катализировать превращение субстрата, поэтому скорость ферментативной реакции замедляется. Чем выше концентрация ингибитора, тем больше молекул фермента в каждый момент времени связаны с ингибитором и тем сильнее происходит ингибирование. Поскольку субстрат и ингибитор – структурные аналоги, то в ходе реакций между ними происходит конкуренция за связывание с активным центром фермента, поэтому степень игибирования ферментативного превращения зависит от соотношения концентраций субстрата и ингибитора. Если концентрация субстрата во много раз превышает концентрацию ингибитора, то в каждый момент времени лишь небольшое число молекул фермента связано с ингибитором и действие ингибитора почти не проявляется.
Следует отметить, что конкурентные ингибиторы не являются полными структурными аналогами субстрата, так как для связывания с ферментным белком ингибитору вполне достаточно, если он будет структурно совместим хотя бы с одним из участков связывания субстрата в активном центре фермента, а другая часть молекулы ингибитора может существенно отличаться от молекулы субстрата.
Хорошо изученный пример конкурентного ингибирования – действие малоновой кислоты на фермент сукцинатдегидрогеназу, катализирующий отщепление водорода от янтарной кислоты в цикле ди- и трикарбоновых кислот:
СООН
СН2СООН сукцинатдегидрогеназа СНСООН ½
½ ¾¾¾¾¾¾¾¾¾® || СН2
СН2СООН СНСООН ½
СООН
янтарная кислота фумаровая кислота малоновая кислота
Малоновая кислота – структурный аналог субстрата - янтарной кислоты, поэтому способна связываться вместо субстрата с активным центром сукцинатдегидрогеназы, но подвергаться окислению с образованием фумаровой кислоты малоновая кислота не может, вследствие чего молекулы фермента, связанные с малоновой кислотой, не участвуют в каталитическом действии и скорость превращения янтарной кислоты в фумаровую понижается. Однако если в реакционную среду добавить большое количество субстрата, то есть янтарной кислоты, то ингибирование фермента прекращается.
Фермент рибулозодифосфаткарбоксилаза, катализирующий присоединение СО2 к первичному акцептору в цикле Кальвина (см. стр…), подвергается конкурентному ингибированию повышенной концентрацией О2. Кислород представляет собой структурный аналог СО2, поэтому может связываться с активным центром рибулозодифосфаткарбоксилазы и действует в этом случае как конкурентный ингибитор.
На основе конкурентного ингибирования производится подбор и синтез многих лекарственных препаратов, действие которых направлено на подавление активности ферментов инфекционной микрофлоры. Использование химических препаратов в качестве ингибиторов ферментных систем возбудителей различных заболеваний получило название химиотерапии. Для химиотерапии подбирают такие конкурентные ингибиторы, которые по структуре наиболее полно соответствуют участкам связывания субстрата в активном центре ферментов и поэтому обладают очень сильным ингибиторным действием.
Неконкурентные ингибиторы не имеют структурной аналогии с субстратами и поэтому взаимодействуют не с участком связывания субстрата в активном центре фермента, а с другим участком ферментной молекулы, вызывая инактивацию каталитического центра. При этом ингибитор не вступает в конкурентное взаимодействие с субстратом и не препятствует связыванию субстрата в активном центре фермента, однако молекула субстрата не подвергается превращению, так как под влиянием ингибитора становятся неактивными группировки фермента, которые активируют субстрат. Поскольку при неконкурентном ингибировании субстрат и ингибитор связываются с разными участками молекулы фермента, действие такого ингибитора не ослабляется при увеличении концентрации субстрата в физиологической среде.
По механизму неконкурентного ингибирования действуют цианиды, которые подавляют активность ферментов, содержащих атомы Fe или Cu (цитохромоксидазы, фенолоксидазы и др.). Ингибитор, содержащий цианидные группировки (CN‾), образует комплексное соединение с металлом, входящим в состав фермента, в результате чего атом металла становится не способным участвовать в каталитическом действии фермента и скорость ферментативной реакции понижается. А в состоянии насыщения фермента ингибитором она практически полностью прекращается. При удалении цианида из реакционной среды каталитическая активность фермента восстанавливается. Неконкурентное ингибирование могут вызывать также катионы водорода, которые вследствие малых размеров не препятствуют связыванию субстрата, но при соединении с некоторыми группировками в активном центре фермента изменяют их электрический заряд и тем самым инактивируют каталитический центр.
В процессе необратимого ингибирования молекула ингибитора образует прочную ковалентную связь с одной из группировок в активном центре фермента, в результате чего становится невозможным его каталитическое действие. В связи с тем, что образовавшееся соединение ингибитора с ферментом не разрушается и не диссоциирует в условиях физиологической среды, в которой функционирует фермент, то каталитическая активность фермента подавляется необратимо.
Многие ингибиторы, вызывающие необратимое ингибирование, обладают строго избирательным действием, так как способны взаимодействовать только с определенными группировками в активном центре фермента. Так, например, многие галогенопроизводные органических веществ являются специфическими ингибиторами ферментов, содержащих в актив-ном центре сульфгидрильные группы (HS – группы). Взаимодействие таких ингибиторов с молекулой фермента происходит по следующей схеме:
Фермент–SH + X–R ¾® фермент–S–R + HX,
где Х – атомы галогенов (Cl, I, Br, F).
Фосфорорганические соединения необратимо ингибируют ферменты, имеющие в активном центре гидроксильные группы аминокислоты серина. Взаимодействие одного из таких ингибиторов диизопропилфторфосфата с ферментом можно представить в виде следующей реакции:
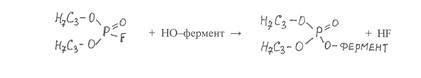 |
В результате присоединения к молекуле фермента фосфорорганического радикала происходит блокирование активного центра фермента и очень сильное подавление его каталитической активности.
Фосфорорганические ингибиторы – сильные яды, необратимо ингибирующие ферменты, которые катализируют гидролиз сложных эфиров холина (холинэстеразы), а также белков (сериновые протеиназы). Одним из ферментов, чувствительных к указанным ингибиторам, является ацетилхолинэстераза, которая катализирует гидролиз ацетилхолина, образующегося в клетках нервных тканей при передаче нервных импульсов. В результате блокирования активного центра ацетилхолинэстеразы фосфорорганическим ингибитором прекращается гидролиз ацетилхолина и он накапливается в нервных клетках, нарушая передачу нервных импульсов, что вызывает паралич и смерть организма. Такие ингибиторы используют в сельском хозяйстве в качестве инсектицидов нервно-паралитического действия.
Все ферменты необратимо ингибируются катионами тяжелых металлов (Hg2+, Pb2+, Ag+, и мышьяка As+), а также галогенопроизводными уксусной кислоты (трихлоруксусная, иодуксусная кислоты и др.), которые при соединении с сульфгидрильными группами (-SH) ферментного белка образуют нерастворимые соединения. Следует отметить, что все факторы, вызывающие денатурацию белков, неспецифически подавляют действие любого фермента, так как основу его составляет молекула белка.
В семенах, клубнях, листьях и других органах растений выявлены специфические белки с молекулярными массами от 5000 до 60000, которые образуют с ферментами неактивные комплексы, в результате чего происходит ингибирование ферментативных реакций. Такие белки называют белковыми ингибиторами ферментов. В зависимости от особеннос-тей взаимодействия с ферментами различают белковые ингибиторы эндогенного и экзогенного действия.
Белковые ингибиторы эндогенные действия образуют неактивные комплексы с ферментами собственного организма и таким образом участвуют в регулировании определенных биохимических процессов в тканях и органах растений. Так, например, в процессе созревания зерновок злаковых растений в них усиливается синтез белковых ингибиторов амилаз и протеаз, катализирующих соответственно гидролиз крахмала и запасных белков, вследствие чего к концу созревания зерновок большая часть указанных ферментов связывается с белками – ингибиторами. Благодаря такому действию ингибиторов происходит накопление в зерне крахмала и запасных белков.
Белковые ингибиторы экзогенного действия подавляют каталитическую активность ферментов чужеродного организма. Выяснено, что некоторые растительные белки образуют неактивные комплексы с пищеварительными ферментами насекомых, животных и птиц, вызывая задержку их роста. К таким ингибиторам относятся некоторые альбумины, содержащиеся в зерновках злаковых и семенах бобовых растений. Они специфически подавляют активность a-амилаз, трипсина и химотрипсина. А белок зерна пшеницы пуротионин обладает бактерицидным и фунгицидным действием.
|
|
|
|
|
Дата добавления: 2014-01-11; Просмотров: 873; Нарушение авторских прав?; Мы поможем в написании вашей работы!