КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Эволюциясы. Өсімдіктер дҮниесініҢ шыҒу тегі мен
|
|
|
|
Тарау
ӨСІМДІКТЕР ДҮНИЕСІНІҢ ШЫҒУ ТЕГІ МЕН
Өсімдіктердің эволюциясының негізгі кезеңдері
Өсімдіктер дүниесінің шығу тегі мен эволюциясын жердің даму тарихымен байланыстырып қарастырған жөн. Жердің даму тарихын әдетте бірінен соң бірі келіп отыратын алты геологиялық эра түрінде береді. Бұлар катархей, архей, протерозой, палезой, мезозой және кайнозой эралары. Оларды өз кезегінде геологиялық кезеңдерге(дәуірлерге) бөледі. Әрбір эраның өзінің шөгінді пародалары болады, олардан өсімдіктердің тасқа айналған қалдықтары мен тастың бетіне түскен іздерін табады. Осы қалдықтарды зерттеу органикалық дүниенің эволюциясының жалпы көрінісін
(бағытын), ал кейде жойылып кеткен өсімдіктерді қалпына келтіруге (реставрировать) мүмкіндік береді.
Катархей эрасында (4,5млрд. жыл бұрын) әлемдік мұхиттарда физико-химиялық процестер жүрген. Олар тіршіліктің пайда болуына қажетті жағдай(субстрат пен материал)туғызған.
Біздің планетамызда тіршілік архей эрасында(2,7-4,5млрд. Жыл бұрын)әлемдік мұхитта пайда болған. Алғашқы өсімдіктердің қашан пайда болғандығын дәл кесіп айту, әзірге мүмкін болмай отыр. Ертедегі өсімдіктердің денесі аса қарапайым іркілдек плазмалардан тұрған. Соған байланысты олардың тасқа айналған қалдықтары біздің уақытымызға дейін сақталмай, із түссіз жоқ болып кеткен. Бірақта олардың тіршілік әрекеттерінің өнімдері, бактериялардан және балдылардан пайда болған әртүрлі пародалар түрінде, сонымен бірге алғашқы организмдердің белоктік денесінің құрамына кірген аминқышқылының қалдығы түрінде сақталған. Бұлар тіршіліктің ең қарапайым формалары- клеткалық құрылысы қалыптаспаған, бір клеткалы ядросы қалыптаспаған, бір клеткалы ядролы және колониялы организмдер болған (259-сурет).
Жер бетінде тіршіліктің дамуы, әсіресе бүкіл геологиялық жылсанаулардың жартысынан көбісін түзетін архей және протерозой эраларында өте баяу жүрген. Мысалы, палеонтологтардың мәліметтері бойынша, тек протерозой эрасында

250-сурет. Тіршіліктің эволюциясы:
1-19-қазіргі кездегі флораның өкілдері; 1- фагтар; 2 –вирустар; 3-бактериялар; 4-5-көк-жасыл балдыралар; 6,9,11,12-жасыл балдырлар; 7-эвгленалы балдырлар; 8-диадомды балдырлар; 10-сары-жасыл балдырлар; 13-саңырауқұлатар; 14-қыналар; 15-псилот; 16-қырықбуын тәрізділер; 17-папоротник тәрізділер; 18-ашық тұқымдылар; 19-жабық тұқымдылар.
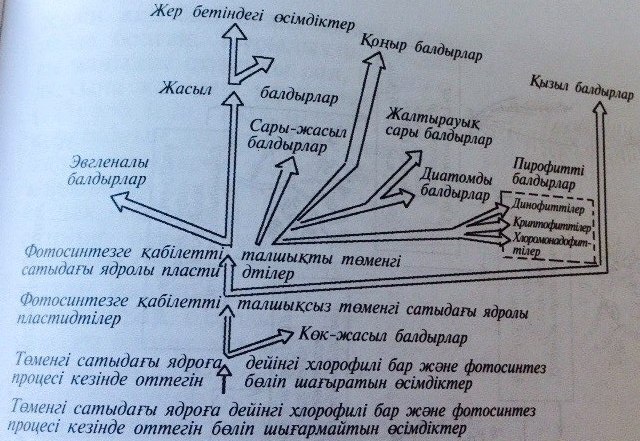
260-сурет. Балдырлардың шығу тегі мен эволюциясы.
(570-2700млн. жыл бұрын)қоректенуі жағынан бір- бірінен айырмашылықтары бар, тірі табиғаттың екі негізгі бағыты анықталды: автотровты(өсімдіктер дүниесі) және гетеротрофты (жануарлар дүниесі) организмдер. Осы эрада бір клеткалы және колониялы көк –жасыл балдырлармен бірге қызыл және жасыл балдырлар (260-сурет), бактериялардың жаңа топтары мысалы темір бактериясы пайда болған.
Өсімдіктер дүниесінің даму тарихында аса маңызды жаңалық палеозой эрасының силурий және девон дәуірлерінде болды (230-570 млн. жыл бұрын). Силурий дәуірінде (405-440 млн. жыл бұрын) риниофиттер- алғашқы құрлықта өмір сүрген өсімдіктер пайда болған (261-сурет.) Бұлар әлі үлкен өсімдіктер болмаған (биіктігі 15-20см-ден аспаған). Олар көп жағдайда құрлықты клем секілді тұтас жауып жатқан. Шамасы осы кезде саңырауқұлақтар, мүктер, қыналар өмір сүрген. Девон дәуірінде (360-405 млн. жыл бұрын) жоғары сатыдағы өсімдіктер жер бетіне түгелдей тарала бастаған. Олар үлкен мөлшерге дейін жеткен. Көптеген өсімдіктердің өмірлік циклінде ұрпақ алмасуы (гаметофазасы мен спорофазасы) қалыптасқан. Бұл процесс өсімдіктерге өте пайдалы болған, өйткені олар өмірінің бір бөлігін сулы ортада (гаметоза), ал екінші бөлігін құрлықта (спорофаза) өткізуге мүмкіндік алған. Спорофиттің құрлықта өмір сүруіне байланысты өсімдік
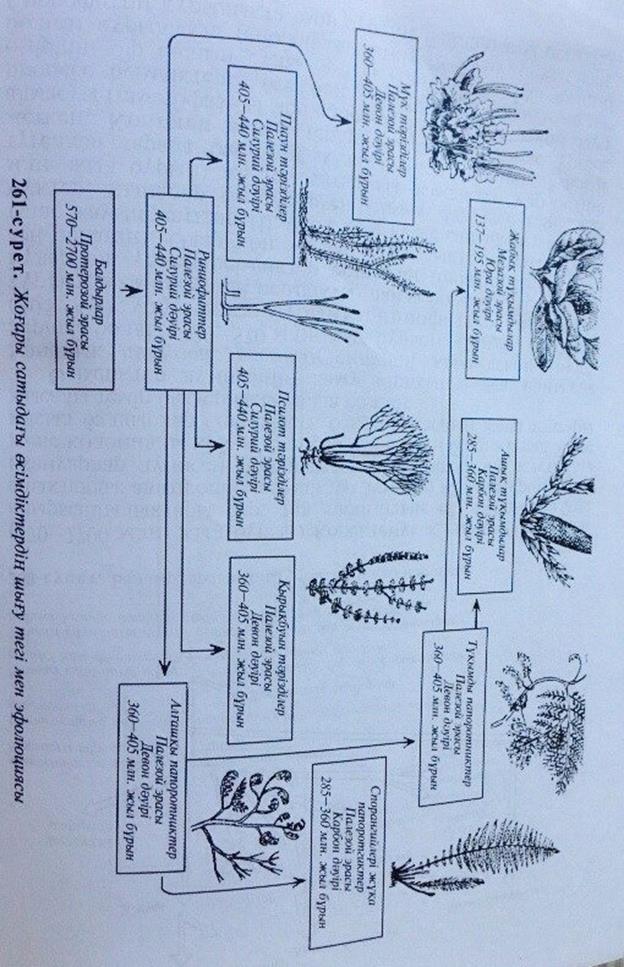 тердің жер беті бөлігі жақсы дамыған, өткізгіш шоқтары пайда болған. Құрлықта өсімдіктердің эволюциясының ең маңызды стимулы сол, олардың табиғи жолмен будандасуына кеңінен мүмкіндік ашылған. Таскөмір дәуірінде (285-350 млн. жыл бұрын) тұқымды папоротниктер мен қырықбуын тәрізділердің өкілдері- каламиттер, қылқан жапырақшаларға жақын биіктігі 30-м-ге дейін баратын кордаиттар кеңінен таралып, өсе бастаған. Қай жерде ұйық батпақ, бойына су сіңген топырақ болса, сол жерде бұл ағаштар орман түзген. Мұндай ормандар теңіз қолтықтарының (лагуна) жағалауларында аз болмаған. Су шайып кеткен жерлерде, әсіресе су тасқындары кезіңде, ағаштар құлап қирап, олардың үстін ұйық басып отырған. Осындай ұйықты батпақтың көп мөлшерде жиналып қалың, шөгінді түзуіне байланысты оттегі жетіспей ағаштар көп мөлшерде шіріп таскөмірдің кеніне айналған (залежи каменного угля). Таскөмір дәуірінде риниофиттер жойылып кеткен.
тердің жер беті бөлігі жақсы дамыған, өткізгіш шоқтары пайда болған. Құрлықта өсімдіктердің эволюциясының ең маңызды стимулы сол, олардың табиғи жолмен будандасуына кеңінен мүмкіндік ашылған. Таскөмір дәуірінде (285-350 млн. жыл бұрын) тұқымды папоротниктер мен қырықбуын тәрізділердің өкілдері- каламиттер, қылқан жапырақшаларға жақын биіктігі 30-м-ге дейін баратын кордаиттар кеңінен таралып, өсе бастаған. Қай жерде ұйық батпақ, бойына су сіңген топырақ болса, сол жерде бұл ағаштар орман түзген. Мұндай ормандар теңіз қолтықтарының (лагуна) жағалауларында аз болмаған. Су шайып кеткен жерлерде, әсіресе су тасқындары кезіңде, ағаштар құлап қирап, олардың үстін ұйық басып отырған. Осындай ұйықты батпақтың көп мөлшерде жиналып қалың, шөгінді түзуіне байланысты оттегі жетіспей ағаштар көп мөлшерде шіріп таскөмірдің кеніне айналған (залежи каменного угля). Таскөмір дәуірінде риниофиттер жойылып кеткен.
Осы бір аса алыс кезеңдерде құрлықтағы жарық пен жылудың ара салмағы бірдей болған. Өйткені құрлықтың осы аздаған беті шексіз жылы акваториямен шайылып отырған. Оны көптеген арнасы суға толы үлкен өзендер мен батпақты кеңістіктер қиып өтіп отырған. Құрлықтың үстіне жердің ішкі жылуы да эффективті түрде әсер етіп отырған. Сонымен бірге, ауаның ылғалдығы да аса жоғары болған, бұлттар жердің бетін қалың болып қаптап, жауып тұрған. Осылардың барлығы құрлықтың климатын теңелуіне әкеліп соқтырған.
Перм дәуірінде (230-285 млн. жыл бұрын) тұқымды папоротниктер және кардаидтер жойылған, қырықбуын тәрізділер мен плаунтәрізділер азая бастаған. Бірақ ашық тұқымдалардың жаңа түрлері, оның ішінде саговниктер, саговниктердің арғы тегі болып табылатын араукариялар және басқа да қылқан жапырақтылар мен гинкголор кеңінен тарай бастаған. Перм дәуірінің соңына қарай ашық тұқымдылар жер бетінде басқа өсімдіктерден басым болған. Мезазой эрасының (67-230 млн. жыл бұрын) триас (195-230млн. жыл бұрын), негізінен юра дәуірлерінде (137-195млн. жыл бұрын) ашық тұқымдылардың флорасы бұрынғысынан әлде қайда артып, молая түсті. Юра дәуірінің ормандары негізінен саговниктерден, гинкголордан, кейтониялар пайда болған. Соңғыларында нағыз жеміс жапырақшалардың гомологы мен шырынды жемістер болған.
Бор дәуірінде (67-137млн. жыл бұрын) жабық тұқымды өсімдіктер кеңінен тараған. Бұлардың пайда болуы өсімдіктер дүниесі мен жануарлар дүниесінің тарихындағы ең шешуші кезең болды. Жабық тұқымдылардың ішіндегі ағаштары магнолия, лавр, платон, эвкалипт, фикус, терек, тал, бук, жүзім және т.б. жер бетінде үлкен кеңістіктерді баса бастады. Осы кезде ағаш тәрізді дара жарнақтылар – пальмалар пайда болып жер бетіне тарала бастады. Шөптесін өсімдіктерден жаңа ортаға бейімделушіліктің негізінде суда өсетін формалар ерекше көріне бастады.
Кайнезой эрасында (0-67 млн.жыл бұрын) жабық тұқымдылар ерекше қарқынмен дамыды. Бұл эраның екі жоғарғы кезеңінде палноген (67-25 млн.жыл бұрын) мен неогенде (25-0,7 млн. жыл бұрын) аса жылы климат қалыптасқан. Сол кездің өзінде- ақ жер бетінің ландшафты қазіргі кездегі жақын болған. Жалпақ жапырақты ормандар пайда бола бастаған. Ағаштардың ішінде нан ағашы (хлебное дерево) инжир, мирт, емен, үйеңкі, шаған, грек жаңғағы және т.б. кеңінен таралған өсімдіктердің тізімін толықтыра түсті. Қылқанжапырақтылардың таза біртектес ормандар түзуде де, аралас ормандар түзуде де (жалпақ жапырақты ағаштармен қосылып) маңызы орасан зор болды. Аса бай шөптесіп өсімдіктер флорасы қалыптасты.
Келесі – төртінші (осы кезгі) кезең 0,7 млн. жылғка созылады. Бұл кезеңде жер бетіндегіклиматта күрт өзгерістер болды: температура төмендеп, ауадан түсетін ылғалдың (жаңбыр, қар) мөлшері арта түсті. Мұның өзі жоғарғы ендікте жекелей алғанда, солтүстік ендікте аса көп мөлшерде мұздың қалыптасуына әкеліп соқтырады. Бұл үлкен мұз оңтүстікке қарай ығысқан. Сонымен мұз дәуірі басталды. Мұздың ең көп басқан уақытында, мұз Европаның орталық аудандарына дейін жеткен. Қазіргі кездегі Украинаның территориясында мұз Днепрдің бойымен Днепропетровск және Житомир облыстарының территорияларына дейін жеткен. Осыған дәуірінің соңы адамның пайда болуымен және дамуымен тікелей байланысты. Адамның пайда болуы өсімдіктер жабынының құрамының өзгеруіне әкеліп соқтыратын жаңа фактор болып табылады. Осы кезден бастап пайдалы өсімдіктерді мәдени жағдайға ендіру қолға алынды, нәтижесінде жабайы өсімдіктер ығыстырыла бастады.
Сонымен, өсімдіктердің даму тарихын үш кезеңге бөлуге болады: балдырлық кезең (протерозой эрасы – палезой эрасының бірінші жартысы), жоғарғы споралы өсімдіктер кезеңі (палезой эрасының екінші жартысы), тұқымды өсімдіктер кезеңі- ашық тұқымдылар (мезеазой эрасының екінші жартысынан осы кезге дейін) пайда болған кезеңдері.
АЛҒАШҚЫ ҚҰРҒАҚҚА ШЫҚҚАН ӨСІМДІКТЕР
Палентологияның мәліметтері бойынша, құрғаққа шыққан алғашқы өсімдіктер риниофиттер болған, олар осыдан 400 млн. жылдар бұрын пайда болған, шамасы жоғарғы деңгейде жетілген хара балдырлары типтес өсімдіктер болса керек. Риниофиттер негізінен батпақты жерлерде өсетін өсімдіктер болған. Олардың вегетативтік денесі телом “ телом” деп аталынады және ол тек қазба түрінде табылған өсімдіктерге ғана тән. Риниофиттердің вегетативтік денесінің өркенге дейінгі құрылысы, дихотомиялық бұтақтанудан, бүйірлік моноподияальды бұтақтанып өсуге ауысуды көзге оңай елестетуге мүмкіндік береді. Ол жоғары сатыдағы өсімдіктердің көптеген топтарында айқын байқалады. (171-сурет)
Теломның жер асты бөлігі бір клеткалы ризоиды бар тамырсабаққа ұқсас болады.
Риниофиттер басқа жоғары сатыдағы өсімдіктерге бастама берген (261-сурет). Мүк тәрізділердің шығу тегі аяғына дейін толық дәлелденбеген. Ғалымдардың бір тобы оларды риниофиттерден шығарады; ал екінші бір тобы- жасыл балдырлардан шығарады. Соңғы көзқарас біршама көңілге қонарлық. Қалай болғанда да мүк тәрізділер эволюциясының тұйық бұтағы.
АЛҒАШҚЫ ЖАБЫҚ ТҰҚЫМДЫЛАР
Жабық тұқымдылардың шығу тегі және олардың мүмкін болған арғы тегі ғалымдарды ылғи да ойға қалдырап келеді. Бұл мәселенің дұрыс шешілуіне осы ғасырдың бас кезінен бастап көп күш жұмсалып келеді. Жабық тұқымды өсімдіктерге көңілдің көп бөлінуі, олардың табиғаттағы және халық шаруашылықтағы маңызының ерекше зор болуымен тікелей байланысты. Оған мынадай бір мысал келтірсек жеткілікті болады. Жер бетінде жабық тұқымды өсімдіктер пайда болған кезден бастап, сүт қоректі жануарлардың және адамның дамуына бірден- бір қажетті мүмкіндіктер туды. Өйткені жабық тұқымды өсімдіктер пайда болғанға дейін, жер бетінде сүт қоректілер жейтін қорек болмаған.
Әдетте жабық тұқымдылардың шығу тегін, гүлдің шығу тегімен байланыстырады. Гүлдің шығу тегі жөнінде бірнеше теориялар бар. Соңғы уақыттарға дейін ең дәлелденген және жалғыз шамамен дұрыс деп табылған эванттық теория болған. Ол стробилярлық теориямен қосылып, гүлді ашық тұқымдылардың стробилінен (шишкасынан) шығарады. Алайда соңғы кездерде ғалымдардың көпшілігі гүлдің шығуының неміс ғалымы В.Циммерман ұсынған теломдық теориясын көбірек мойындап жүр. Бұл мойындап жүр. Бұл теория бойынша алғашқы жоғары сатыдағы өсімдіктер, риниофиттер болған, олардан папоротниктер бастау алған, соңғыларынан өз кезегінде ашық тұқымдылар мен жабық тұқымдылар пайда болған. В.Циммерманның (1930-1965) пайымдауы бойынша жоғарғы сатыдағы өсімдіктердің барлық органдары қарапайым органнан – теломнан пайда болған. Эволюциялық процестің барысында теломдар бірігіп стерильді синтеломдар жапыраққа және сабақтың өсіне бөлінеді, ал фертильді теломдар спорофиллдерге айналады. В.Циммерманның пікірінше гүлдің пайда болу процесімен қатар (параллель) жүрген. Мұндай түсіндірмеден жапырақ пен сабақтың эволюциялық тұрғыдан қарағанда принциптік айырмашылығының болмайтындығын байқау қиын емес. Шындығында жапырақты және өстік органдары (вегетативтік және генеративтік органдарға дифферециацияланбаған) теломдардың системасынантарихи дамудың барысында пайда болған деп қарастырған жөн.
Осы айтылғандардан, жабық тұқымдылардың арғы тегі тек ашық тұқымды өсімдіктер болуы еш талас туғызбайды. Жабық тұқымдылардың мүмкін болған арғы тегі ретінде ашық тұқымдылардың белгілі қатарларының барлығы ұсынылды. Бірақ олардың кейбір мүмкін деп алынған алғашқы формалары вегетативтік денесінің микроскопиялық құрылысының (гнетопсидтерде түтіктердің болуы) жабық тұқымдылардың қарапайым топтарына (түтіктері болмайды)
Қарағанда жоғары деңгейде жақсы жетілген. Сол себептенде бұл формалардан жабық тұқымдыларды шығарудың мүмкіндігі жоқ. Егерде жабық тұқымдылардың мүмкін болған арғы тегін анықтауда олардың сүрегінің структурасына шешуші орын берсек, онда жабық тұқымдылардың арғы тегі қарапайым саговниктер немесе беннеттиттер болуға тиісті. Бірақта мұндай жауапкершілігі жоғары тұжырымға өсімдіктің тек микроскопиялық құрылысының белгілерімен шектелу әрине жеткіліксіз. Сондықтанда салыстырып қарауға және талдау жасауға өсімдіктің жалпы құрылысының көптеген тұрақты белгілерін, мысалы репродуктивтік органдарының құрылысының белгілерін алу қажет. Осыған байланысты ашық тұқымдылардың стробилдеріне (шишки), спорангилеріне және тұқымбүрлеріне салыстырмалы түрде жан- жақты талдаулар жасалынды. Көптеген және әртүрлі салыстырмалы морфологиялық мәліметтерді жинақтай келе,систематик –филогенетиктердің көпшілігі мынадай тұжырымға келген. Қазіргі кездегі белгілі ашық тұқымдылардың ішінен, жабық тұқымдылардың арғы тегі не тұқымды папоротниктер, не болмаса беннеттиттер болуға тиісті. Сонымен бірге мынадай қызық жорамал айтылған. Беннеттиттер мен жабық тұқымдылардың арғы тегі олардың екеуіне де ортақ бір өсімдік болуы мүмкін. Жабық тұқымдылардың гүлінің пайда болуына бенеттиттердің қалайда қатысының барлығы дәлелдене түсті. Қос жынысты гүлдің алғашқы моделін тек қос жынысты стробилден (шишкадан)шығаруға болады (гүлдің шығуының стробилярлық теориясы),ал мұндай стробил (шишка) қазіргі кезде белгілі барлық ашық тұқымдылардың ішінде тек бенеттиттерде ғана бар. Сонымен, классикалық филогенетикалық системаның көптеген көрнекті өкілдері мынадай тұжырымға келген, жабық тұқымдылардың тұқымды папоротниктермен де, саговниктермен де беннеттитермен де туыстық байланысты болған.Бірақта жабық тұқымдылардың шығу тегі туралы гипотезалардың ешқайсысыда мынадай “таңқаларлық фактіні ” түсіндіре алмайды. Ч. Дарвиннің сөзі бойынша жабық тұқымдылардың солтүстік ендікте көп мөлшерде және түрлерінің саны жағынан алуан түрлі болып, барлық континенттерде бір мезгілде пайда болуын. Сонымен бірге олардың алғашқы рет қай жерде пайда болғандығы да белгісіз болып отыр.
1954 жылы белгілі – географ және систематик-эволюционист М.Г.Попов жабық тұқымдылардың эволюциясы жөнінде бұрынғы айтылып жүрген теориялардан принциптік айырмашылығы бар жаңа теория ұсынды. Жабық тұқымды өсімдіктердің шығу тегі мен эволюциясының проблемаларын шешуге жол салған бұл теорияға, бор дәуірінің бас кезінде басталған, туыстығы жағынан бір- бірінен алыс жатқан түрлер мен туыстардың арасында жүрген буындасу процесстері негіз болды. Осы теорияны дәлелдеу мақсатында автор мынадай постулатқа сүйенген. Өсімдіктердің жаңа сорттарын шығаруда, біздің уақытымызда үлкен масштабта жүріп жатқан будандастыру процесі, ертедегі геологиялық эпохаларда, түрлердің түзілуінде бұданда үлкен роль атқарған. Архегониялы өсімдіктердің систематикалық топтарына морфологиялық талдаулар жасаудың негізінде М.Г.Попов мынадай қорытындыға келген. Архегониялы өсімдіктердің вегетативтік денесінің структурасында үш түрлі негізгі тип айқын басымдық көрсетеді: филлофитті, бұл жағдайда жапырақтардың системасы басым болып келеді, сабақтары не бұтақтанбайды, не болмаса нашар бұтақтанады, метамериясы қалыптаспаған (папоротник тәрізділер,бенеттиттер); артрофитті немесе бунақты, бұл жағдайда скелетті өстердің системасы басым болып келеді, өркеннің метамерлері айқын байқалады (қырықбуын тәрізділер,гнетопсидтер); микрофилді –ұсақ жапырақты, бұл жағдайда әртүрлі систематикалық топтардың арасында байланыстар не үзілген, немесе нашар қалыптасқан болып келеді (плаун тәрізділер). Осыған байланысты жабық тұқымдыларды берген алғашқы аталық және аналық өсімдіктер (родительские пары) М.Г.Поповтың жорамалдауы бойынша бенеттиттер мен гнетопсидтер болған. Осындай алыс будандасудың негізінде, аталық пен аналықтың көптеген белгілері бір мезгілде ажыраған. Соған байланысты жабық тұқымдылардың тұқымдастарының типінің тұтастай “веер” секілді шашыраған тармақтары қалыптасқан. Олардың вегетативтік денесінің структурасында аталық және аналық өсімдіктердің екеуініңде белгілері болған.
Алғашқы тұқымдастық топтарды ағаштардың түрлері түзген, сондықтанда эволюция зоналар бойынша, негізінен саматикалық тұрғыдан жүрген: яғни дамудың бағыты ағаштардан, шөптесін өсімдіктерге қарай жүрген. Соңғылары екі бағытта дамыған: биік жоғарғы ендікте суыққа төзімді, немесе криофильді өсімдіктер пайда болған (көп жылдық шөптесін өсімдіктер), ал оңтүстікке қарай шөлейт айтмақтарда-құрғақшылыққа төзімді, немесе аридті өсімдіктер пайда болған(бір жылдық өсімдіктер).
Жабық тұқымдылардың шығу тегінің “будандасу процесі теориясы” гүлді өсімдіктердің бірден тропикалық зонаның әртүрлі аудандарында түзелгендігін және көптеген тұқымдастардың типінің кенеттен пайда болғандығының себептерін түсіндіреді. Сонымен бірге алғашқы жабық тұқымдастырдың қай геологиялық эпохада пайда болғандығы мен шығу тегін анықтауға және тағы басқа көптеген сұрақтарға жауап беруге мүмкіндік береді. Бұл теория филогенетикалық системаны түзуге бірден- бір жол ашты. М.Г. Поповтың теориясы эволюциялық теорияны, тың ойлардың жаңа қырларымен толықтырды. Табиғат туралы ғылымның фундаментальдық проблемаларын, тіптен кеше ғана шешімі табылмайды деп келген мәселелерді шешудің жаңа жолдарына бағдар береді. Жабық тұқымдылардың мәңгілік жұмбақ сырларын шешуде, бірден- біртығыз байланыста болатын, жақын ғалымдардың, оның ішінде генетиканың күштерін біріктіре отырып, одан әрі батыл қадам жасау, болашақта қандай ғылымның жолдарының айқын шындыққа әкелетіндігін көрсетеді.
ӨСІМДІКТЕРДІҢ ЭВОЛЮЦИЯСЫНЫҢ ЖАЛПЫ
ЗАҢДЫЛЫҚТАРЫ
КӨБЕЮДІҢ ЭВОЛЮЦИЯСЫ
Көбею тірі табиғаттың ерекше қасиеттерінің бірі ретінде, оның дамуының ең алғашқы кезеңдерінде пайда болған. Өсімдіктердің эволюциясымен бірге тіршіліктің бастапқы формаларынан бастап қазіргі кезге дейін көбеюдің жолдары да белгілі бір эволюциялық кезеңдерден өтіп, түрлене өзгерген. Сонымен бірге көбеюдің эволюциясының ең маңызды қозғаушы күші болады және жаңа арнайы маманданған органдардың түзілуіне әкеліп соқтырады.
Жыныссыз көбею – ең ерте пайда болған және күмәнсіз көбеюдің бастапқы тәсілі болып табылады. Ол өсімдіктердің барлық топтарына, соның ішінде жабық тұқымдыларға да тән.
Жыныссыз көбеюдің ең қарапайым тәсілі вегетативтік жолмен көбею. Оны клеткалық құрылысы қалыптаспаған организмдердің өмірінің әртүрлі кезеңдерінен кездестіруге болады. Көптеген ядросы қалыптаспаған организмдерде (мысалы, көк жасыл балдырларда бұл тәсіл, қазіргі кездің өзінде көбеюдің жалғыз жолы болып табылады. Көп жағдайда өсімдіктердің әрбір табиғи топтарының өздеріне тән вегетативтік көбеюдің формалары болады. Мысалы, көк – жасыл балдырларда бұл гормониялар, қыналарда-издиилер мен соредиилер, саңырауқұлақтарда- оидиилер мен телиоспоролар, мүк тәрізділердің бірқатарында – өнім бүршікшелері (133,168,147,174-суреттер).Жоғарғы сатыдағы өсімдіктерде, әсіресе жабық тұқымдыларда вегетативтік көбеюдің формалары алуан түрлі болып келеді.
Нағыз жыныссыз көбеюге арналған споролардың түрлері көп болады. Эволюцияның барысында олар аналық өсімдікке спорофитке ұқсас жаңа особьтарды түзу қасиетінен айрылған, бірақ жыныстық ұрпаққа (гаметофитке) бастама береді.
Жыныстық көбею эволюциясының өте ертедегі кезеңдерінде пайда болған, бірақ оның пайда болған, бірақ оның пайда болған уақытын дәл кесіп айту қиын. Өсімдіктердің қазіргі кездегі кейбір қарапайым топтарының жыныстық процесі белгісіз. Сонымен көк жасыл балдырлар секілді кейбір табиғи топтарда жыныстық көбеюдің еш уақытта болмағаны күмән келтірмейді. Соңғы уақыттарға дейін дробянкаларды да осы топқа жатқызып келген. Алайда дробянкаларды электронды микроскопты пайдалана отырып зерттеудің нәтижесінде, оларда жыныстық көбеюдің капуляциялық, трансформациялық және трансдукциялық түрлерінің болатындығы анықталды. Басқалары, мысалы кейбір саңырауқұлақтар, шамасы жыныстық жолмен көбею мүмкіндігінен айрылған. Жыныстық жолмен көбею мүмкіндігін жоғалту фактасы, кейбір жоғары деңгейде жетілген өсімдіктерде де, мысалы жабық тұқымдыларда белгілі (партогенез).Әзірше ғылымда, өсімдіктердің жыныстық процесінің эволюциясының толық картинасын көрсететіндей мәліметтер жеткіліксіз. Алайда эволюцияның төменгі деңгейінде тұрған өсімдіктердің эволюциясының жоғарғы деңгейінде тұрған өсімдіктерге қарағанда жыныстық процесі өте қарапайым формада өтетіндігі толық дәлелденген. Эволюцияның деңгейінің бірден бір көрсеткіші, ол гаметалардың мамандануы. Ең қарапайымдысы изогомиялық жыныстық процесс. Біршама жоғарғы деңгейде жетілген жыныстық процеске гетерогамия жатады. Ең жоғары деңгейде жетілген жыныстық процеске оогамия жатады (130-сурет). Жер бетінің жағдайында тіршілік етуге бейімделуіне байланысты аталық гамета қозғалту қабілетінен айырылады және жұмыртқа клеткасына тозаң түтігі арқылы жеткізіледі.
Сонымен, тұқымды өсімдіктерде жыныстық процесс сулы ортамен байланысты емес, ол дегеніміз өсімдіктердің бұл тобының құрлықта өмір сүруіне бейімдеогендігі болып табылады.
Жыныстық көбею жыныссыз көбеюден бөлек емес, қайта керісінше онымен байланыста дамымаған. Өсімдіктердің әрбір табиғи топтарының өмірлік циклінде, жыныссыз және жыныстық көбеюлердің ауысып келуінің өзіндік заңдылықтары болады. Төменгі сатыдағы өсімдіктердің көпшілігінде ядролық фазалардың алмасуы болмайды, өмірлік циклден спорофитті түгелдей шығып қалған деседе болғандай (тек зигота ғана диплоидты). Кейбір төменгі сатыдағы өсімдіктердің және барлық жоғарғы сатыдағы өсімдіктердің өмірлік циклінде
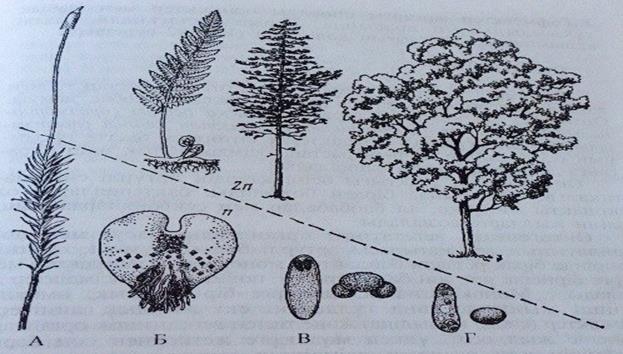
262-сурет. Жоғары сатыдағы өсімдіктердің әртүрлі систематтикалық топтарының спорофиті мен гаметофитінің ара қатынасы:
А-Мүк тәрізділер; Б-папоротник тәрізділер; В-жалаңаш тұқымдылар; Г- жабық тұқымдылар.
спорофит пен гаметофит алмасып келіп отырады.Олардың морфологиялық әртектілігінің деңгейі және өсімдіктердің зерттеп отырған тобының өмірлік циклінде басым болып келуі эволюциясының деңгейінің маңызды көрсеткіші болып таблады. В.Гофмейстердің (1849, 1851ж.ж.) жұмыстарының негізінде аса маңызды жалпы заңдылық ашылған: эволюциянын барысында спорофит бірітндеп жақсы дамиды және дербестікке ие болады; гаметофит, керісінше, редукцияға көбірек ұшырап, өзінің дербестігін толығымен жоғалтады және спорофитке түгелімен бағынышты болады. Бұл жағдай жоғары деңгейде дамыған өсімдіктерде - ащық тұқымдылар мен жабық тұқымдыларда байқалады (262-сурет). Өсімдіктердің әртүрлі табиғи топтарының өмірлік циклін зерттей отырып,В. Гофмейстер жоғарғы споралы өсімдіктер мен жабық тұқымдылардың арасында, бұрын ойлағандай үлкен алшақтықтың жоқтығын дәлелдеді (180,192-суреттер).
|
|
|
|
|
Дата добавления: 2014-11-06; Просмотров: 11867; Нарушение авторских прав?; Мы поможем в написании вашей работы!