КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Загальні відомості
|
|
|
|
Тема: Відділ Голонасінні, соснові (Pinophyta).
Лабораторна робота №7
Мета: Ознайомитись із характерними ознаками класів голонасінних. На прикладі окремих представників розгялнути цикл розвитку голонасінних.
До заняття підготувати наступні питання:
1. Місце голонасінних в системі органічного світу.
2. Ознаки ксероморфності голонасінних.
3. Характерні ознаки класу насінні папороті (Pteridospermae).
4. Характерні ознаки класу саговникові (Cycadopsida).
5. Характерні ознаки класу бенетитові (Bennettitopsida).
6. Характерні ознаки класу гінкгові (Ginkgopsida).
7. Характерні ознаки класу гнетові (Gnetopsida).
8.Характерні ознаки класу хвойні (Pinopsida).
9. Цикл розвитку сосни звичайної.
10. Особливості чоловічого гаметофіту сосни звичайної.
11. Розвиток жіночого гаметофіту сосни звичайної.
12. Значення голонасінних у природі та житті людини.
13. Місце теми в шкільному курсі біології.
Голонасінні, соснові (Pinophyta)
Сучасні голонасінні - це невелика група, блиько 800 видів рослин, поширених на всй земній кулі, але більшість їх розселена вздовж узбережжя Тихого океану, в Австралії і на прилеглих островах. Група дуже давня. Виникли голонасінні в палеозойській ері як закономірний етап в еволюції вищих спорових рослин, якому передували кілька важливих подій: виникнення різноспоровості (яка була відома ще з середнього девонського періоду), а також камбію і деревних форм. Таким чином, передумови появи голонасінних уже були підготовлені всім ходом еволюції. Найкращим підтвердженням цього була знахідка у верхньодевонських відкладах Пінічної Америки насінини, яку назвали архоспермою.Це найдавніша насінина, вік її 370 млн. років.
Таким чином, предків голонасінних слід шукати серед палеозойських рослин і особливо різноспорових, найдавніших форм папоротеподібних.
Дрібнолиста (мікрофільна) гілка еволюції сучасних голонасінних виникла, можливо, від кордаїтів. Останні походять від спільних з насінними папоротями давніх різноспорових папоротеподібних. Кордаїти найбільш вірогідно дали початок гінкговим і хвойним. З ним гінкгові зближує характер розміщення насінних зачатків і внутрішня будова стебла, а з хвойними - будова стробілів, анатомічна будова хвої.
Основна відмінність голонасінних від інших архегоніальних рослин - наявність насінини як одиниці розселення. Від спорофіта відокремлюється не спора, а багатоклітинне утворення - насінина, що містить зародок майбутнього спорофіта і запас поживних речовин. Насінина утворюється з насінного зачатка, видозміненого в процесі еволюції мегаспорангія, що розвивається відкрито на насінних лусках жіночої шишечки (стобіла). Звідси і назва цієї групи рослин - голонасінні.
Всі голонасіні - різноспорові, одно- або дводомні рослини. Мікроспори розвиваються в мікроспорангіях, які утворюються на мікрофілх чоловічої шишечки. Мегаспори розвиваються в насінних зачатках.
Голонассіним також властива зміна поколінь і ядерних фаз, але порівняно з папоротеподібними статеве покоління дуже спрощене. Чоловічий гаметофіт розвивається в середині мікроспори і складається з кількох клітин. Він зовсім позбавдений антеридіїв, у більшості голонасінних чоловічі гамети не мають джгутиків і перетворені у спермії. Жіночий гаметофіт розвивається у нуцелусі насінного зачатка (мегаспорангії), ніколи не контактує з навколишнім середовищем, тому голонасінні належать до ендопроталіальних (внутрішньозаросткових) рослин.
Гаметофіти живуть 1-3 роки. Запліднення здійснюється в середині нуцелуса і не залежить від наявності води.
Незалежно від води запліднення, захищеність зародка майбутього спорофіта, наявність насінини - великий крок вперед на шляху еволюції рослинного світу.
Спорофіти голонасінних досить розвинені, це дерева, кущі з головним коренем, моноподіальним галуженням, наявністю камбію, більш досконалою провідною системою, довговічні.
Завдання 1. Ознайомитись із класифікацією та характерними ознаками класів голонасінних.
Голонасінні діляться на шість класів: насінні папороті, саговникові, або цикадові, бенетитові, гінкгові, гнетові, хвойні,або шишконосні.
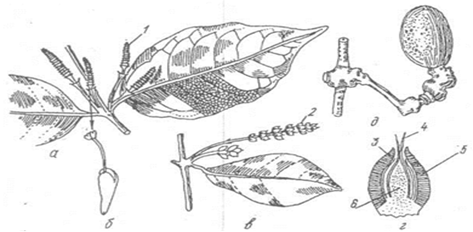
Клас насінні папороті (Pterydospermae). Найдавніші викопні рослини, що жили, починаючи з верхнього девонського до кінця тріасового періоду, найбільшого розквіту досягли в кам'яновугільний і пермський періоди. Вперше їх викопні рештки були описані у 1904 р. англійськими палеоботаніками Ф.Олівером і Д.Скоттом. Це були високі дерева або ліани з великими пірчастими листками, розміщено спірально. Розмножувалися насінням, в якому не були знайдені зародки. Насінні зачатки розвивались на кінцях бічних, верхівкових сегметнів листків. На верхівці насінного зачатка містися жіночий гаметофіт (ендосперм), оточений нуцелусом. На гаметофіті містились архегонії з яйцеклітинами (мал.29).
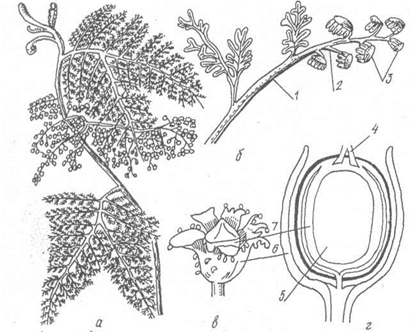
Мал.29 Насінна папороть: а - загальний вигляд (фрагмент рослини), б - репродуктивний пагін, в - насінний зачаток, вкритий мисочкою, г - поздовжній розріз насінного зачатка з мисочкою: 1 - вегетатична частина пагона, 2 - спороносна частина пагона, 3 - мікроспорофіли з мікроспорангіями, 4 - мікропіле, 5 - нуцелус, 6 - мисочка, 7 - інтегумент.
Мікроспорангії розміщувались по краю мікроспорофіліві заповнювались численними мікроспорами. Внутрішня будова стебла досить складна: у центра знаходилась добре розвинена серцевина, оточена колатеральними пучками. У пучку, крім камбію, були вторинні елементи флоеми і ксилеми, ксилема складалась тільки з трахеїд. Ззовні стебло вкривала масивна кірка.
Представниками цього класу є роди: лігіноптерис, каліматотека, гросотека та ін.
Філогенетично насінні папороті вважаються перехідною групою між папоротеподібними і насінними рослинами.
Клас саговникові, або цикадові (Cycadopsida). Це сучасні та викопні рослини, загальна їх кількість тепер становить близько 120-130 родів, поширених у тропічних і субтропічних зонах Східної Азії, Австралії, Африки і Америки.
Клас об'єднує одну родину і 9 родів. Це переважно дерева з колоноподібними стовбурами і великими пірчастими листками. Тому систематики минулого, серед них і К.Лінней, відносили їх до пальм. Вирішальну роль у визначенні їх місця в систематиці відіграли праці відомого німецького ботаніка В.Гофмейстра (1824-1877), який зруйнував панівне на той час уявлення про відсутність філогенетичних зв'язків між папоротеподібними і голонасінними рослинами.
Саговники - роздільностатеві рослини. Чоловічі шишечки поодинокі, розміщуються на верхівках стовбурів, їх довжина від 2 до 50-80 см. Мікроспорофіли несуть на нижньому боці групи мікроспорангіїв, заповнених дрібним пилком. Пилок має дві оболонки: зовнішню товстішу - екзину і внутрішню тоншу - інтину. Ядро мікроспори ділиться, з нього утворюється спочатку три ядра, а потім і три клітини, з яких одна найдрібніша є, по суті, єдиною вегетативною клтиною чоловічого гаметофіта. Друга клітина є антеридіальною, вона дає початок двом великим сперматизоїдам з численними джгутиками. Третя клітина згодом, під час запліднення, перетворюється у пилкову трубку.
Жіночі шишечки великі, 2-100 см завдовжки, несуть мегаспорофіли різної форми, на яких утворюється по кілька пар насінних зачатків. Насінні зачатки 3-5 см завдовжки мають тришаровий інтегумент, зовнішній і внутрішній - м'ясистий, середній - твердий. Під внутрішнім інтегументом знаходиться нуцелус, в якому з єдиної мегаспори розвивається жіночий гаметофіт з кількома архегоніями, що розміщуються у пилковій камері (мал.30). Пилок, що потрапляє на насінний зачаток, проростає, пилкова трубка вростає в нуцелус і доносить до рідини пилкової камери сперматизоїди, які там плавають певний час. Потім один із сперматизоїдів запліднює яйцеклітину і зигота дає початок зародку, а насінний зачаток перетворюється у насінину, інтегументи розростаються, утворюючи м'ясисту шкірочку насінини.
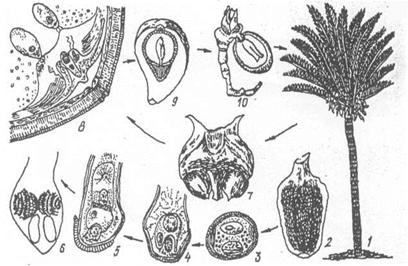
Мал.30. Цикл розвитку саговника: 1 - загальний вигляд, 2 - мікроспорофіл, 3-6 розвиток чоловічого гаметофіта з мікроспори, 7 - мегаспорофіл у цератозамії, 8 - верхівка насінного зачатка саговника, 9 - насінина в розрізі, 10 - проросла насінина.
Із цього класу найпоширенішими родинами є роди замія і саговник (цикас). Практичне значення невелике, їх розводять як декоративні рослини, а із серцевини стовбура одержують крохмаль і готують сагову крупу. Саговники цікаві насамперед як реліктові рослини, що збереглися з давно минулих часів, і свідчать про походження голонасінних.
Клас бенетитові (Bennettitopsida). Це велика група палеозойських рослин, що почали своє існування в тріасовому і вимерли в крейдяному періоді. Ці рослини мали прості, галузисті, товсті чи тонкі стебла, за зовнішнім виглядом нагадували як насінні папороті, так і саговники. Листки пірчасті, рідше цілісні, прості. За анатомічною будовою стебла також близькі до саговників. У центрі стебла розміщувалась серцевина із смоляними каналами, оточена первинною і вторинною ксилемою, що складалась лише з трахеїд. Між ксилемою і флоемою було камбіальне кільце.
Характерною рисою бенетитів була двостатева ентомфільна шишка (стробіл). Шишка мала потовщену вісь конічної форми, на ній при основі знаходились стерильні пірчасті листочки, вище розміщувались пірчасті мікроспорофіли, на яких сиділи зрослі у синангії мікроспорангії. В центрі стробіла розташовувались мегаспорофіли, які несли по одному насінному зачатку (мал.31). Насінний зачаток складався з інтегументів, нуцелуса і жіночого гаметофіта. Насіння бенетитів мало шкірочку, ендосперм і зародок з двома м'ясистими сім'ядолями.

Мал.31. Вільямсонія: а - загальний вигляд, б - двостатева шишка в розрізі, в - поздовжній розріз через мегаспорангії, г - поздовжній розріз насінини (1 - зародок, 2 - нуцелус, 3 - лусочка), д - поздовжній розріз насінного зачатка.
Через наявність двостатевого стробіла деякі вчені розглядали бенетитові як проміжну групу між голонасінними і квітковими рослинами.Так, англійські палеоботаніки В.Арбер і Д.Паркін висловили гіпотезу, що стробіл бенетитових є прототипом квітки магнолієвих, це твердження відобразилось у створеній авторами стробілярної теорії походження квітки. Цим і вичерпується філогенетичне значення цього класу.
Клас гінкгові (Ginkgopsida). Єдиним сучасним представнико цього класу є реліктовий вид - гінкго дволопатеве, рослина з родини гінкгових. У минулому представники родини жили у верхньому тріасовому періоді, розквіту досягли у юрському періоді мезозойської ери. Для науки цю рослину відкрили вперше у 1712 році. В природному стані вона росте на півдні Китаю, але з давніх давен її широко культивують як ритуальну рослину в різних районах Східної Азії. В Європу завезена у XVIII ст., де її розводять у ботанічних садах як релікт.
Спорофіт гінкго - високе дерево з гладкою ясно-сірою корою і пірамідальною кроною. Листки з віялоподібною пластинкою, розміщені поодиноко на видовжених гонах і групами - на вкороченнях. Стебло має добре розвинену ксилему, складену трахеїдами, тонкий шар кори, камбій і серцевину. У різних частинах стебла, кореня і листків є секреторні клітини і лізигенні смоляні канали.
Гінкго дволопатеве - дводомна рослина. Чоловічі стробіли сережкоподібні, кожен мікроспорофіл складається з нжки і двох спорангіїв, наповнених пилком. Пилок триклітинний. Кожен мегаспорофіл складається з ніжки і двох спорангіїв, наповнених пилком. Пилок триклітинний. Кожен мегаспорофіл складається з насінного зачатка і особливого валика при основі (мал.31), вони не зібрані у стробіли і утворюються на кінцях вкорочених пагонів у пазухах листків. Насінний зачаток має інтегумент, витягнений на вершку у пилковхід (мікропіле), нуцелус і жіночий гаметофіт з архегоніями. Після злиття одного з багатоджгутикових сперматизоїдів з яйцеклітиною розвивається зародок з двома сім'ядолями, оточений масивним ендоспермом (залишки жіночого гаметофіта). Насіння гінкго їстівне, багато на олію, крохмаль і ароматичні сполуки (мал.32).
Гінкго дволопатеве має наукове значення як реліктова рослина з архаїчними ознаками організації: дихотомічним жилкуванням листків, багатоджгутиковими сперматизоїдами, соковитою оболонкою насіння. Практичне значення невелике, розводять як декоративну рослину.
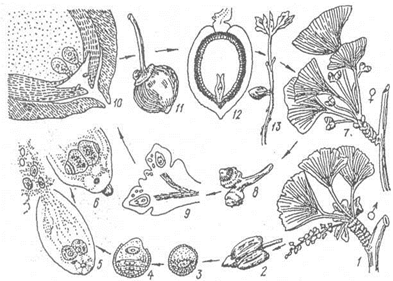
Мал.32. Цикл розвитку гінкго дволопатевого: 1 - гілка з мікроспорофілами, 2 - мікроспорофіл, 3-6 - розвиток мікроспори, 7 - гілка з мегаспорангіями, 8 - мегаспорангій, 9 - мегаспорангій у розрізі, 10 - верхівка насінного зачатка, 11 - насінина збоку, 12 - насінина в розрізі, 13 - проросла насінина.
Клас Хвойні, або Пінопсиди (Pinopsida). Це центральний і найчисленіший клас серед сучасних голонасінних, який об'єднує близько 600 видів, 55 родів і 8 родин.
У викопному стані хвойні відомо з верхнього карбону, а розквіт їх припадає на мезозойську еру. У палеогені кайнозойської ери вони були представлені майже сучасними родами, але мали ширший ареал, який скоротився у неоні та антропогені внаслідок різкої зміни клімату.
Хвойні відіграють велику ландшафтно-утворювальну роль, бо 90% лісів земної кулі складається саме з них, або хвойні входять до них як складова частина. Ці ліси мають рекреаційне, водо- і грунтозахисне, санітарно-гігієнічне, естетичне значеня, а також дають цінну деревину, що має широке і цінне застосування.
Усі сучасні хвойні - це дерева або кущі, деякі з них є гігантами рослинного світу, наприклад секвойя, що сягає до 120 м заввишки і до 16 м у діаметра, живе до 3000 років і більше.
Галуження більшості з них моноподіальне, стовбури стрункі, вкриті кіркою, деревина має чітко виражені річні кільця і складається переважно з трахеїд, пронизана вертикальними і горизонтальними смоляними каналами, що містять ефірні олії, смоли. Коренева система стрижнева, на молодих корінцях розвивається мікориза. Листки вузькі (це мікрофільна гілка еволюції голонасінних), живуть 2-11 років, мезофіл однорідний, кутикула масивна, епідерміс товстостінний, під ним добре розвинена гіподерма, глибоко занурені продихи, в центрі розміщений провідний циліндр (жилка). Це типова ксероморфна будова листка, яка пов'язана з недосконалістю провідної системи стебла.
Клас хвойних поділяється на два підкласи: кордаїтиди і хвойні.
Підклас Кордаїтиди - це викопні форми, що були поширені на початку мезозойської ери. Це були стрункі дерева з голчастими листками і дрібними шишечками. Їх залишки утворили великі поклади кам'яного вугілля.
Підклас Хвойні. Він охоплює сім родин, з яких дві - вольцієві і подозамітові - викопні рослини, коротка характеристика інших наводиться нижче. Серед хвойних найдавнішими є родини араукарієві і подокарпові.
Родина араукарієві (Araucariaceae). Включає два роди і 40 видів, поширених у Південній півкулі. Це одна з родин серед сучасних хвойних із цілим рядом примітивних ознак. За антомічною будовою деревини вони близькі до кордаїтів. Це дво-, рідше однодомні рослини з високим стовбуром. Листки широкі, рідше дрібні, голчасті. Чоловічі і жіночі шишечки великі, від 4 до 30 см завдовжки. Рослини з роду араукарія часто утворюють ліси, наприклад араукарія бразильська – одна з основних лісоутворювальних порід в Андах. Араукарію високу, що росте в Австралії, у нас культивують як оранжерейну рослину. З рослин роду агатис одержують цінну деревину і дамарову смолу.
Родина подокарпові (Podocarpaceae). Об'єднує 8 родів і 130 видів, поширених переважно у поза тропічних областях Південної кулі, де вони є головним лісоутворювальними породами. Найважливіші роди – подокарпус і дакридіум. У межах природного ареалу вирощують як деревні рослини, а в субтропіках Кавказу і Криму – як декоративні рослини.
Родина тисові (Taxaceae). Об'єднує 2 роди і 20 видів, поширених у Північній півкулі. Дерева і кущі з плоскою хвоєю. Мікроспорофіли щиткоподібні, зібрані у короткі шишечки. Мегаспорофіли не зібрані у шишечки, мають по 1-2 насінних зачатки, які розташовані в пазухах лускуватих листків біля верхівки пагонів. Після запліднення насінного зачатка утворюється насінина з твердою кам'янистою оболонкою і яскравим принасінником (арилюсом). У тисових деревина без смоляних каналів, тверда, стійка проти гниття. Тому великі тисові гаї, які були на Північному Кавказі, в Криму, Карпатах, вирубані ще у минулому столітті. Ареал тисових різко скоротився, а культивування їх пов'язане з великими труднощами, рослини повільнорослі, тіньовитривалі, вибагливі до вологості грунту і повітря. У гірському Криму, Карпатах і Прикарпатті подекуди збереглися зарості тису ягідного (Taxus baccata) занесено до Червоної книги України.
Родина соснові (Pinaceae). Об'єднує 10 родів і близько 250 видів, поширених переважно у північній півкулі, де утворюють обширні ліси. Соснові однодомні, рідше дводомні дерева або кущі. Листки (хвоя) лінійні, голчасті, розміщені спірально, поодиноко, попарно, групами (на укорочених погонах). У більшості видів листки вічнозелені, тільки у модрин опадні.
Чоловічі шишечки зібрано групами, жіночі поодинокі, різної будови та забарвлення. Пилок, звичайно, з двома повітряними мішечками. Розглянемо найбільш поширені роди.
Рід сосна (Pinus) - найчисленніший, налічує 100 видів. Це переважно високі дерева, що мають вкорочені й видовжені пагони. Хвоя розміщена на вкорочених пагонах пучками, по 2-3-5 хвоїнок. У природному стані зростають тільки дво- і п'ятихвойні сосни: сосна звичайна (P.sylvestris), гірська (P.mugo), кримська (P.pallasiana) - двохвойні, а сибірська (P.sibrica), або кедр сибірський, як її частіше називають - п'ятихвойна.
Сосна звичайна -це одна з найважливіших лісоутворювальних порід в Україні, вона формує ліси на піщаних і супіщаних грунтах, це так звані бори, які відіграють важливу грунтозахисну роль. Сосна звичайна дає цінну деревину, є джерелом одержання живиці.
Сосна сибірська - могутнє дерево, поширене по всьому Сибіру. Крім високоякісної деревини, використовується насіння, так звані кедрові горішки, які вживають у їжу, з них одержують олію. В Карпатах зрідка трапляється сосна європейська, яку розводять також як декоративну рослину. До складу соснових лісів гірського Криму входить сосна Палласова, або кримська, на мисі Піцунда зберігся реліктовий і ендемічний вид - сосна піцундська (P.pithyusa).
Рід модрина (Larix) нараховує близько 20 видів, з них в Україні - один вид, а в культурі успішно випробувано 10 видів. Модрини - це високі стрункі дерева з опадними. Модрини - стрункі дерева з опадними хвоїнками, що сидять на укорочених пагонах по 2-40-60 шт. Швидкорослі, світлолюбні рослини. Шишечки дрібні, насіння дозріває протягом року. Деревина міцна, стійка проти гниття, багата на смолу. У формуванні хвойних лісів модринам належить провідне місце, особливо у північно-східних районах Європи і Азії. Найбільш поширеними у культурі України є модрина сибірська (L.sibrica) і європейська (L.decidua).
Рід ялина (Picea) об'єднує 40-45 видів, поширених у холодних і помірних областях Північної півкулі. В Україні зростає два види. Ялини - це високі дерева з пірамідальною кроною і дрібними загостреними на верхівці хвоїнками, що тримаються 10-12 років. Зрілі шишечки циліндричні, пониклі, довго висять на деревах після висипання насіння, яке дозріває протягом року. Найважливішим представником є ялина звичайна (P.abies), яка утворює чисті або мішані ліси, на великих просторах, дає високоякісну деревину, придатну для виготовлення паперу, музичних інструментів, в Карпатах – гірська (P.montana). Рослини декоративні, придатні для поодиноких посадок, створення алей. Особливо декоративні голуба і срібляста форми ялини колючої (P.pungens), що походять в Північної Америки.
Рід ялиця (Abies) нараховує 45 видів, у флорі України – один. Великі дерева із кільчастим розміщенням гілок, хвоя плоска з двома продиховими смужками білого кольору з нижнього боку. Жіночі шишечки прямостоячі, після дозрівання насіння розсипаються. У флорі Росії найпоширенішою є ялиця сибірська (A.sibrica), яка утворює ліси на великих просторах. У Карпатах поширена ялиця біла, або європейська (A.alba). Дає м'яку деревину, що використовується як будівельний матеріал, придатна для виготовлення паперу, хімічної переробки. Із хвої одержують ефірну олію, з живиці – так званий страсбурзький терпентин. Ялиці – декоративні рослини.
Рід кедр (Cedrus) об'єднує 4 види. В Україні не росте. Це красиві могутні дерева, які утворювали великі гаї у Гімалаях, на Близькому Сході, Малій Азії, Північній Америці. Особливо відомі кедр гімалайський (C. deodara), кедр ліванський (С.libani). Здавна культивуються в садах і парках Європи.
Родина таксодієві (Taxodiaceae). Об'єднує 10 родів і 15 видів, поширених переважно у Північній півкулі. В Україні представників родини немає. До цієї родини належать дерева-велетні, що живуть понад 3-4 тис.років. Це секвойя червона, поширена на західному узбережжі Північної Америки; секвойядендрон гігантський (Segujoiandendron giganteum), дерево, що досягає до 120 м заввишки і до 16 м у діаметрі. Ці рослини взято під охорону.У нас їх розводять як декоративні рослини. До родини належать також і метасеквойя (Metaseguoia glyptostroboides) – рослина, яка вважалась вимерлою. У 1946 р. китайські ботаніки несподівано виявили зарості цієї рослини і з того часу її розводять як релікт палеогену в садах і парках різних країн світу.
Родина кипарисові (Cupressaceae). Об'єднує 20 родів і близько 150 видів, поширених на всій земній кулі, у флорі України – один вид. Це одно-, дводомні кущисті, деревоподібні рослини. Листки дрібні колючі, лускоподібні. Жіночі шишечки невеликі, після дозрівання дерев'яніють чи стають м'ясистими. Насінні і покривні луски зростаються, утворюючи луску, що несе 1-2 чи багато насінних зачатків.
Рід кипарис (Cupressus) об'єднує 15 видів, поширених у помірно теплих областях Європи, Азії, Північної Америки. У нас культивують 4 види в Криму. Найпоширенішим є кипарис вічнозелений – Cupressus sempervirens.
Рід яловець (Juniperus) нараховує 70 видів, поширених в північній півкулі, у флорі України є 8 видів. Це кущі, невеличкі дерева з лускоподібними, голчастими листками, розміщеними по три в кільцях. Луски жіночих шишечок м'ясисті, при дозріванні стають соковитими, утворюючи так звану шишкоягіду. Насіння дозріває протягом двох років. Усі ялівці світлолюбні, невибагливі до родючості грунту рослини, що ростуть на рівнині й піднімаються високо в гори, іноді досягають крайньої межі поширення деревних форм. У нашій флорі поширені такі види, як яловець звичайний (J.communis), козацький (J.sabina), високий (J.exselsa). Ялівці – декоративні рослини, деревина йде на виготовлення дрібних виробів, деякі види отруйні.
З інших видів часто культивують як декоративні рослини тую західну (Thuja occindentalis), біоту східну (Platycladus orientalis).
Клас гнетові (Gnetopsida), або оболонконасінні (Chlamidospermatopsida). Храктерною рисою оболонконасінних є наявність справжніх судин у вторинній деревині, немає драбинчастих судин, смоляних ходів. Навколо мікроспорофілів і макроспорофілів формуються покриви, які розростаються і огортають насінини своєрідною оболонкою, схожою з оплоднем. Звідси їх назва оболонконасінні. Чоловічі і жіночі гаметофіти дуже редуковані. Зародок з двома сім'ядолями. Вони виникли, можливо, від бенетитових, або мають спільне з ними походження від якихось інших примітивних фори. Схожість з покритонасінними і намагання розглянути їх як перехідні форми немає підстав, вона носить конвергентний характер.
Оболонконасінні діляться на 3 порядки: ефедрові, гнетові, вельвічієві.
Порядок ефедрові (Ephedrales). Включає одну родину ефедрові, монотипний рід ефедра і 40 видів. Це великі кущі з ефільними пагонами.Листки редуковані лускоподібні,супротивні,кільчасті. Функцію фотосинтезу виконують пагони. Продихи розташовані в заглибленнях асиміляційної тканини.
Ефедра - дводомна рослина. На молодих пагонах чоловічих рослин утворюються по 2-8 квіткові шишечки. На осі шишечки в пазухах верхніх лусок розміщені мікроспорофіли. Кожний мікроспорофіл несе 2-8 двогніздових спорангіїів. Нижня частина мікроспорофіла оточена двома лускоподібними листочками або оцвітиною (Мал. 33).
У мікроспорангії утворюється велика кількість мікроспор, які починають проростати ще в мікроспорангії. Спочатку виникає проталіальна клітина, а при наступних поділах формуються вегетативне і антеридіальне ядра. Внаслідок поділу з антеридіального виникає спермагенне ядро і ядро клітини-ніжки. В такому вигляді пилок потрапляє в насінний зачаток.
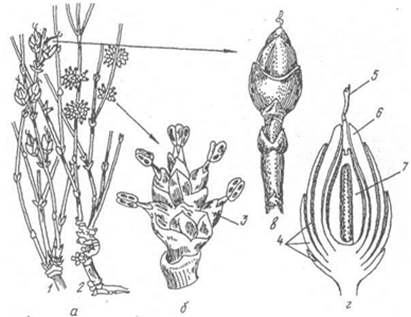
Мал.33. Ефедра двоколоскова: а – загальний вигляд репродуктивних пагонів, б – групи чоловічих шишечок, в – жіноча шишечка, г – розріз жіночої шишечки: 1 – пагін жіночої рослини, 2 – пагін чоловічої рослини, 3 – елементарна чоловіча шишечка, 4 – зовнішній інтегумент, 5,6 – насінний зачаток, 7 – внутрішній інтегумент.
На молодих пагонах жіночих рослин розвиваються жіночі шишечки. Знизу шишечку оточує кілька пар лускоподібних листочків. Закінчується вісь шишечки насінним зачатком. Ззовні насінний зачаток оточує потужний покрив, який нагадує редуковану оцвітину. Внутрішній покрив, або інтегумент витягується в мікрополярну трубку, яка вловлює пилок. Центральну частину виповнює нуцелус, на верхівці якого виникає пилкова камера.
У нуцелусі одна з клітин виділяється своїми розмірами, вмістом поживних речовин і активністю. В результаті її поділу формуються 4 мегаспори, три з них дегенерують, а четверта розвивається в жіночий заросток. На його верхівці формуються два редукованих архегонії. Пилок, потрапивши на такий насінний зачаток, за допомогою мікропілярної трубки досягає пилкової камери і крізь утворену пилкову трубку проникає до нуцелуса і архегонія. Всередині пилку містяться ядра вегетативної і спермагенної клітини. Останнє, ділячись, дає два спермії. Вони потрапляють у архегоній, і один з них зливається з яйцеклітиною, запліднює її. Із зиготи розвивається зародок, що складається із зародкового корінця,ніжки,підсім'ядольного коліна, двох сім'ядолей і брунечки.
Зріла насінина складається із соковитого червонуватого покриву, кам'янистого інтегумента, нуцелуса, ендоспарма і розміщеного в ньому зародка.
Ефедра використовується для одержання ефедрину, що має лікарське значення.
Порядок гнетові (Gnetalis). Цей порядок включає родину гнетових і монотипний рід з 40 видами, поширеними в тропічних вологих лісах Азії, Африми та Америки. Більшість з них ліани і лише два види невисокі дерева. Стебла з супротивними, широкими шкірястими листками. Рослини дводомні. Чоловічі екземпляри мають шишечки у вигляді сережок. На їх осі кільцями по 2-3 розміщені мікроспорофіли. Такий мікроспорофіл в основі оточений покривом оцвітиною, що складається з двох зрослих листочків. Між покривів підноситься сплющена вісь (тичинкова нитка), яка закінчується двома одногніздовими спорангіями. За своєю морфологією мікроспорофіли нагадують тичинки квіткових (мал. 34). В спорангіях визрівають численні мікроспори. Кожна з них проростає в дуже редукований чоловічий гаметофіт - пилок. Чоловічий заросток, що виникає, дуже редукований. Проталіальна клітина не утворюється, є вегетативне ядро, ядро клітини-ніжки і ядро спермагенної клітини, що виникли внаслідок поділу антеридіальної клітини. У такому вигляді пилок потрапляє на насінний зачаток.
Жіночі шишечки таож мають вигляд сережок. На їх осі розміщені кільцями насінні зачатки. Насінний зачаток оточує покрив - оцвітина. Глибше знаходяться інтегументи, зовнішній з яких коротший, а внутрішній витягнутий в мікропілярну трубку. Центральну частину займає нуцелус.
Мал.34. Гнетум гнетоподібний: а – репродуктивний пагін чоловічої рослини, б – елементарна чоловіча шишечка, в – репродуктивний пагін жіночої рослини, г – розріз жіночої шишечки, д – насінина: 1 – група чоловічих шишечок, 2 – група жіночих шишечок, 3 – зовнішній інтегумент, 4 – внутрішній інтегумент, 5 – середній інтегумент, 6 – нуцелус.
В ньому утворюються 4 мегаспори, три з них дегенерують,а четверта розростається в дуже редукований жіночий гаметофіт, який немає архегоніїв і повністю розвиненої тканинної будови. Тільки в нижній частині заросток тканинний, а у верхній містяться лише ядра.
Ще такий насінний зачаток потрапляє пилок, який проростає і утворює пилкову трубку. По ній переміщуються вегетативне ядро і спермагенна клітина, яка ділиться і дає початок двом сперміям. Пилкова трубка досягає гаметофіта і виливає спермії, які зливаються з найближчими вільними ядрами жіночого гаметофіта. З виникаючих зародків повністю розвивається тільки один. Він складається із зародкового корінця, підсім'ядольного коліна, двох сім'ядолей і брунечки.
Насінина складається із зовнішнього соковитого шару, кам'янистого шару (зовнішнього інтегумента), тоненької плівки (внутрішнього інтегумента), маслянистого перисперма і розмішеного в ньому зародка.
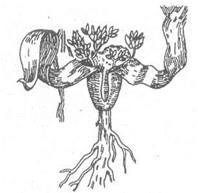
Порядок вельвічієві (Welwitschiales). До них належить родина вельвічієві з родом Welwitschia і одним видом W.mirabilis. Вельвічія – ендемік пустині Наміб. Від кореневої шийки в грунт проникає,
корінь до 1-1,5 м завглибшки. Верхівка стебла сідлоподібна увігнута; від її країв відходять в протилежні боки два шкірясті листки, які протягом ста років досягають 2-3 м завдовжки (мал.35).
Мал.35. Молода рослина вельвічії дивної
Вельвічія - дводомна рослина. На верхівках пагонів чоловічих екземплярів групами зібрані чоловічі шишечки. На спільній осі розміщені криючі листки і мікроспорофіли; останні зібрані групами по шість і зростаються основами в трубочку. Ззовні вони оточені чотирма лусочками (оцвітиною), з яких дві коротші, а дві довші. Мікроспоофіл утворює 3 спорангії. В них формуються численні спори. Проростають вони ще в спорангії. Чоловічий заросток (пилок) дуже редукований і складається із трьох ядер: проталіального, вегетативного і сперматогенного.
На жіночих екземплярах розвиваються групами жіночі шишечки, оточені знизу чотирма безплідними лусочками. В пазухах цих лусочок розміщені насінні зачатки. Кожний насінний зачаток оточують 2 зрослі лусочки (оцвітина). Нерідко ці лусочки ботаніки сприймають за зовнішній інтегумент. Внутрішній інтегумент витягнутий в мікропілярну трубку. Вся внутрішня частина заповнена нуцелусом. У верхній частині його відбувається мегаспорогенез; утворюється чотири мегаспори, з яких три зникають, а четверта дає редукований жіночий заросток.
Архегонії не виникають. Заросток представлений дуже зрослою мегаспорою, в цитоплазмі якої вільно розміщені численні ядра. Потім ядра скупчуються по 2-3 у верхній частині і 10-12 в нижній. Між ними виникають перетинки і заросток набуває клітинної форми. Одноядерні клітини верхньої частини заростка витягуються в протріальні трубки і досягають верхньої частини нуцелуса.
Пилок, потрапляючи на нуцелус, утворює пилкову трубку, яка вростає в тканину нуцелуса і змикається з протріальною трубкою. Спермагенне ядро пилкової трубки ділиться і утворює два спермії. Один з них зливається з ядром проталіальної трубки. Із заплідненого ядра в нижній частині заростка розвивається зародок, який складається із зародкового корінця, ніжки, підсім'ядольного коліна і двох сім'ядолей.
Зріла насінина складається із крилатого зовнішнього покриву, тонкої оболонки (інтегумента), соковитої тканини (перисперми, нуцелуса) і зародка.
Завдання 2. На прикладі сосни звичайної (Pinus sylvestris) розглянути цикл розвитку голонасінних.
Хвойні – одно- і дводомні рослини, спори утворюються в роздільностатевих шишечках (стробілах). Цикл розвитку їх можна простежити на прикладі сосни звичайної (мал.36). Навесні на верхівках торішніх пагонів утворюються групи чоловічих шишечок жовтого кольору. Кожна шишечка довжиною до 10 мм і складається з осі, до якої прикріплюються мікроспорофіли, що несуть два паралельні мікроспорангії. У мікроспорангіях мейозом утворюються мікроспори, які мають дві оболонки – екзину (внутрішню) та два повітряні пухирці, що сприяють кращому розвіюванні мікроспор. Спочатку мікроспора одноклітинна і одноядерна, але ще у мікроспорангії внаслідок поділу ядра утворюються чотири клітини, що є недорозвиненим чоловічим гаметофітом: дві клітини - проталіальні, швидко розсмоктуються, ще одна – антеридіальна і остання, більша – вегетативна. Двоклітинний чоловічий гаметофіт називається пилковим зерном. Остаточний розвиток його відбувається на насінному зачатку.
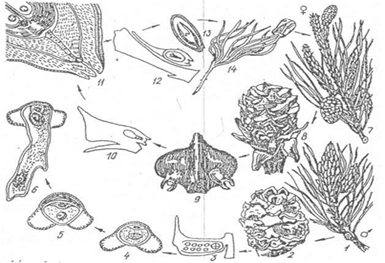
Мал.36. Цикл розвитку сосни звичайної: 1 – гілка з групами чоловічих шишечок, 2 – чоловіча шишечка, 3 - розріз мікроспорангія, 4-6 проростання мікроспори, 7 – гілка з жіночими шишечками, 8 – жіноча шишечка, 9 – насінна лусочка з двома насінними зачатками, 10 – насінна лусочка збоку, 11 – верхівка насінного зачатка, 12,13 – насінина в розрізі, 14 – проросла насінина.
Жіночі шишечки розвиваються на молодих пагонах, вони поодинокі, довжиною до 10 мм і забарвлені у зелений або червонуватий колір. Жіночі шишечки не гомологічні чоловічим, бо будова їх інша. На осі жіночої розміщуються два типи лусочок: покривна і насінна, причому остання розміщується в пазусі покривної. На думку німецького ботаніка А. Брауна (1842), насінна лусочка – короткий пагін з двома листками. Ця думка пізніше пітвердилась дослідженнями ботаніків Л.Челаківського (1900), А.Амса (1913), Р.Флоріна (1954). При основі насінної лусочки є два насінних зачатки. Насінний зачаток – це видозмінений мегаспорангій, він має таку будову: інтегумент (покрив) витягнений на верхівці у мікропіле (пилковхід). Під покривом міститься нуцелус, що зростається з ним. Через певний час одна із клітин нуцелуса, що знаходиться поблизу мікропіле, набуває властивостей архегоніальної (материнської клітини мегаспор). Ця клітина ділиться мейозом на чотири лінійно розміщені мегаспори. Одна з них проростає, а інші поступово дегенерують. Проросла мегаспора перетворюється на багатоклітинний жіночий гаметофіт або інші поступово дегенерують. Проросла мегаспора перетворюється на багатоклітинний жіночий гаметофіт або первинний ендосперм з двома архегоніями спрощеної будови. Пилок вітром переноситься на мікропіле насінного зачатка і проростає. Його вегетативна клітина витягується у пилкову трубку, що вростає в нуцелус і ендосперм. Антеридіальна клітина ділиться ще раз, утворюючи базальну і генеративну клітини. Генеративна клітина, ділячись, утворює два спермії, один з яких запліднює яйцеклітину. Спермії крізь пилкову трубку потрапляють до яйцеклітини, в яку попередньо опускається антеридіальне ядро. Із зиготи утворюється зародок. Часто виникає кілька зародків, це явище називається поліембріонією. Насінний зачаток у процесі розвитку і формування зародка перетворюється у насінину, інтегумент – у шкірку насінини. Сформований зародок має первинний корінець, стебельце і кілька (2-20) сім'ядоль. Первинний ендосперм розростається, витісняючи нуцелус, і використовується як запасні поживні речовини.
Жіночі шишечки після запліднення розростаються, дерев'яніють, після дозрівання насіниння їх лусочки підсихають, розкриваються, насіння висипається і розноситься вітром. Насіння має крилоподібні придатки. У сосни період від запилення до запліднення триває майже три роки.
Література:
1. Григора І.М., Шабарова С.І., Алейніков І.М. Ботаніка К., Фітосоціоцентр. – 2000 р.
2. Комарницкий Н.А., Кудряшов Л.В., Уранов А.А. Систематика растений. М., 1962 р.
3. Нечитайло В.А., Кучерява Л.Ф. Ботаніка. Вищі рослини. К., Фітосоціоцентр, 2000 р.
|
|
|
|
|
Дата добавления: 2014-11-25; Просмотров: 7845; Нарушение авторских прав?; Мы поможем в написании вашей работы!