КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Строение синапса. Синаптическая передача возбуждения ….….. 23 1 страница
|
|
|
|
Клетки, возникающие из нервной трубки, могут быть либо предшественниками нейронов, либо предшественниками клеток нейроглии. Внутренний слой стенки нервной трубки даёт начало эпендиме, выстилающей полости центральной нервной системы, из среднего слоя образуется серое вещество, а из наружного слоя – белое вещество.
Общий принцип развития нервной системы сводится к тому, что возникшие нейроны не остаются на месте появления, а мигрируют на свои окончательные позиции. Миграция обусловлена тем, что нервная система возни- кает в виде тонкой эктодермальной трубки (нервная трубка), а в конечном счёте становится гораздо более крупной структурой – нервной системой.
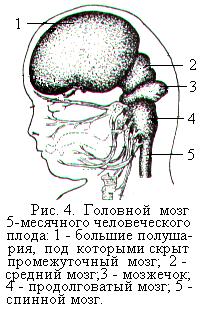
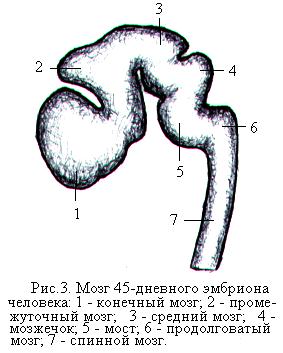 Из нижнего отдела нервной трубки образуется спинной мозг. Головной конец нервной трубки утолщается и уже на 4-й неделе эмбрионального развития различают три мозговых пузыря. К концу 4-й недели передний мозговой пузырь начинает делиться на два, а на 5-й неделе на два делится задний (ромбовидный) пузырь. В результате головной конец нервной трубки состоит из пяти пузырей, из которых развиваются все отделы головного мозга (рис. 3). Из пятого (заднего) мозгового пузыря развивается продолговатый мозг, из четвёртого – варолиев мост и мозжечок, из третьего – средний мозг, из второго – промежуточный мозг, из первого (переднего) – большие полушария головного мозга (рис. 4). Полости мозговых пузырей развиваются в желудочки мозга.
Из нижнего отдела нервной трубки образуется спинной мозг. Головной конец нервной трубки утолщается и уже на 4-й неделе эмбрионального развития различают три мозговых пузыря. К концу 4-й недели передний мозговой пузырь начинает делиться на два, а на 5-й неделе на два делится задний (ромбовидный) пузырь. В результате головной конец нервной трубки состоит из пяти пузырей, из которых развиваются все отделы головного мозга (рис. 3). Из пятого (заднего) мозгового пузыря развивается продолговатый мозг, из четвёртого – варолиев мост и мозжечок, из третьего – средний мозг, из второго – промежуточный мозг, из первого (переднего) – большие полушария головного мозга (рис. 4). Полости мозговых пузырей развиваются в желудочки мозга.
 Наиболее интенсивно растёт первый пузырь. К 6-му месяцу внутриутробного периода большие полушария полностью покрывают мозг. К этому времени все отделы мозга хорошо выражены. До 4-го месяца развития плода поверхность больших полушарий гладкая, на ней отмечается лишь вдавливание будущей боковой борозды. В дальнейшем наружный корковый слой полушарий растёт быстрее внутреннего, что приводит, начиная с 5-го месяца, к появлению борозд, делящих поверхность полушарий на доли, а затем и на извилины.
Наиболее интенсивно растёт первый пузырь. К 6-му месяцу внутриутробного периода большие полушария полностью покрывают мозг. К этому времени все отделы мозга хорошо выражены. До 4-го месяца развития плода поверхность больших полушарий гладкая, на ней отмечается лишь вдавливание будущей боковой борозды. В дальнейшем наружный корковый слой полушарий растёт быстрее внутреннего, что приводит, начиная с 5-го месяца, к появлению борозд, делящих поверхность полушарий на доли, а затем и на извилины.
В процессе развития коры нейроны, мигрировавшие первыми, заполняют сначала глубинные слои, а последующие клетки обосновываются в более поверхностных слоях. Клетки, являющиеся предшественницами нейронов, располагаются в так называемых пролиферативных зонах, выстилающих поверхность желудочков мозга. В этих зонах происходят и все последние митотические деления клеток, становящихся нейронами. Созревание нейронов происходит гораздо медленнее, чем их образование.
У новорожденного ребёнка тела нервных клеток невелики, ветвление дендритов развито слабо. В дальнейшем происходит рост и дифференцировка нервных клеток. В пределах коры раньше всего созревают крупные пирамидные нейроны, а затем более мелкие вставочные нейроны, образующие локальные сети. Созревание нейронов коры связано с формированием нервных сетей коры. В первые дни после рождения ребёнка происходит бурный рост числа синапсов. Одновременно в тех или иных размерах происходят гибель нейронов и перестройка нервных связей.
Все основные борозды и извилины формируются у ребёнка к моменту рождения. Но развитие формы и величины борозд и извилин, формирование мелких новых борозд и извилин продолжается и после рождения. Филогенетически более молодые отделы коры больших полушарий у новорожденных развиты слабее и с возрастом относительно увеличиваются, а более старые, наоборот, с возрастом относительно уменьшаются.
Общий план строения нервной системы
 Нервная система образована нервной тканью, состоящей из нервных клеток – нейронов и клеток нейроглии. Клетки нейроглии выполняют опорную функцию для нервных клеток, входят в состав их оболочек, участвуют в обмене веществ нейронов, выполняют секреторную и защитную функции. В нейроглии различают две группы клеток: 1) макроглию, 2) микроглию. В зависимости от строения и выполняемой функции клетки макроглии в свою очередь подразделяются на эпендимоциты, олигодендроциты и астроциты.
Нервная система образована нервной тканью, состоящей из нервных клеток – нейронов и клеток нейроглии. Клетки нейроглии выполняют опорную функцию для нервных клеток, входят в состав их оболочек, участвуют в обмене веществ нейронов, выполняют секреторную и защитную функции. В нейроглии различают две группы клеток: 1) макроглию, 2) микроглию. В зависимости от строения и выполняемой функции клетки макроглии в свою очередь подразделяются на эпендимоциты, олигодендроциты и астроциты.
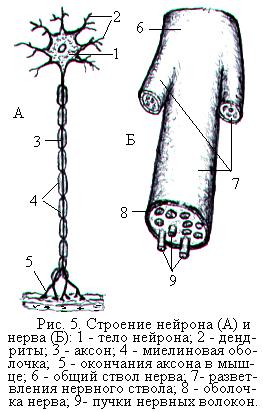
Нейрон является структурно-функциональной единицей нервной системы. Он состоит из тела и отростков (рис. 5, А). Большинство нейронов имеет несколько (от 1 до 1000) коротких ветвящихся отростков – дендритов и один длинный (от нескольких сантиметров до 1,5 м) отросток – аксон. Особенностью нервных клеток является то, что по дендритам импульс может проводиться только к телу нейрона, а по аксону – от тела нейрона.
Отросток нейрона, окружённый оболочками, образованными олигодендроцитами, называется нервным волокном. В зависимости от строения оболочки различают безмякотные нервные волокна, или безмиелиновые, и мякотные нервные волокна, или миелиновые. В безмякотных волокнах олигодендроциты образуют вокруг отростка нейрона однослойную оболочку. В мякотных нервных волокнах плоские отростки олигодендроцитов многократно закручиваются вокруг отростка нейрона, образуя слой миелина. Мякотные волокна значительно толще, чем безмякотные, и проводят нервные импульсы с большей скоростью. Пучки мякотных и безмякотных нервных волокон, покрытые общей соединительнотканной оболочкой, называются нервами (рис. 5, Б).
Нервные волокна, по которым возбуждение (нервный импульс, потенциал действия, биоток) поступает в ЦНС, называются чувствительными, или центростремительными, или афферентными. Нервные волокна, по которым импульсы передаются от ЦНС к рабочим органам, называются двигательными, или центробежными, или эфферентными. В пределах ЦНС, т.е. в спинном и головном мозге, расположены вставочные, или ассоциативные нейроны. По вставочным нейронам возбуждение передаётся с афферентных на эфферентные нейроны, а также распространяется по спинному и головному мозгу. Нервные волокна заканчиваются нервными окончаниями.
 Нервную систему человека условно подразделяют на центральную – ЦНС и периферическую. ЦНС образована спинным и головным мозгом. Головной мозг человека состоит из 6 отделов: продолговатого мозга, варолиева моста, мозжечка, среднего мозга, промежуточного мозга и больших полушарий (рис. 6). Мост и мозжечок часто объединяют под названием задний мозг, а промежуточный мозг и большие полушария – под названием передний мозг. Большие полушария часто называют конечным мозгом. Средний мозг совместно с варолиевым мостом и продолговатым мозгом образует ствол головного мозга. К стволу мозга иногда также относят промежуточный мозг и мозжечок.
Нервную систему человека условно подразделяют на центральную – ЦНС и периферическую. ЦНС образована спинным и головным мозгом. Головной мозг человека состоит из 6 отделов: продолговатого мозга, варолиева моста, мозжечка, среднего мозга, промежуточного мозга и больших полушарий (рис. 6). Мост и мозжечок часто объединяют под названием задний мозг, а промежуточный мозг и большие полушария – под названием передний мозг. Большие полушария часто называют конечным мозгом. Средний мозг совместно с варолиевым мостом и продолговатым мозгом образует ствол головного мозга. К стволу мозга иногда также относят промежуточный мозг и мозжечок.
На разрезе спинного и головного мозга можно заметить тёмные участки – серое вещество мозга и светлые участки – белое вещество мозга. Серое вещество мозга состоит в основном из тел нейронов, а белое вещество – из большого количества нервных волокон и отростков нейронов.
К периферической нервной системе относят многочисленные нервы, отходящие от спинного и головного мозга, нервные узлы и нервные сплетения, расположенные за пределами спинного и головного мозга.
Есть ещё одна классификация нервной системы, согласно которой всю её подразделяют на соматическую и вегетативную, или автономную. Соматическая нервная система иннервирует (снабжает нервами) главным образом стенки тела и конечности, скелетные мышцы, кожу, обеспечивает связь организма с внешней средой. Вегетативная нервная система иннервирует все внутренности, железы, гладкие мышцы, регулирует обмен веществ. Вегетативная нервная система подразделяется на две части: парасимпатическую и симпатическую.
Строение нейрона
Нейрон содержит многие компоненты, присущие всем клеткам живого организма. В нервной клетке выделяют три отдела: тело, или сому, дендриты и аксон. В теле наряду с другими органоидами клетки находятся ядро, рибосомы и происходит синтез необходимых для клетки веществ. Основное отличие ядра нейрона состоит в том, что после рождения человека оно всегда находится в интерфазе, в то время как ядра большинства других клеток организма могут находиться в разных фазах митоза. Интерфазное состояние ядра нейрона сохраняется в течение всей жизни человека, т.к. нервные клетки не делятся.
Нейроны мозга в процессе индивидуального развития человека образуют многочисленные связи между собой. Каждая связь имеет определённое функциональное значение. Это создаёт специфическую для каждого человека индивидуальность ЦНС. Если бы нервные клетки делились в течение всей жизни человека, то образовавшиеся структурные связи между нейронами разрывались бы, постоянно нарушалась бы индивидуальность нервной системы и всё время менялось бы поведение человека. Однако нейроны могут начать делиться при некоторых заболеваниях нервной системы.
Хотя ядра нейронов не делятся и тела их не восстанавливаются, однако отростки нервных клеток могут восстанавливаться после повреждений или заболеваний. Благодаря этому возобновляется чувствительность и подвижность органов после хирургических операций на них.
Несмотря на то, что в течение индивидуальной жизни масса нейронов погибает в результате каких-то нарушений или старения, мозг, тем не менее, сохраняет свои функции. Сохранение функций обеспечивается избыточностью нервных клеток в мозге, многократным дублированием функций, а также перераспределением функций между отдельными нейронами. Даже при обширных поражениях мозга в результате кровоизлияния или хирургического вмешательства люди могут сохранять интеллект и высокую работоспособность.
Нервные клетки в зависимости от их функций делятся на чувствительные, вставочные и двигательные. Чувствительными называют нейроны, которые имеют рецепторные окончания, преобразующие энергию внешнего воздействия в нервный импульс. К двигательным относятся нейроны, аксоны которых идут к исполнительным органам – к мышцам или железам. Больше всего в мозге вставочных нейронов, которые осуществляют связь между чувствительными и двигательными нейронами, а также между различными отделами спинного и головного мозга.
 Нейроны, вызывающие возбуждение в иннервируемых клетках, называют возбуждающими, а приводящие к торможению – тормозными. В основе химической классификации нервных клеток лежат вещества, которые синтезируются в теле нейрона и выделяются его нервными окончаниями. Например, нейроны спинного мозга, управляющие работой скелетных мышц (мотонейроны), выделяют ацетилхолин и называются холинергическими. Некоторые нейроны продолговатого мозга и варолиева моста синтезируют норадреналин и их относят к норадренергическим. В продолговатом мозге имеются также серотонинергические нейроны, секретирующие серотонин, в мозжечке – ГАМК-ергические, которые выделяют гамма-аминомасляную кислоту (ГАМК). В среднем мозге находятся дофаминергические нейроны, выделяющие дофамин.
Нейроны, вызывающие возбуждение в иннервируемых клетках, называют возбуждающими, а приводящие к торможению – тормозными. В основе химической классификации нервных клеток лежат вещества, которые синтезируются в теле нейрона и выделяются его нервными окончаниями. Например, нейроны спинного мозга, управляющие работой скелетных мышц (мотонейроны), выделяют ацетилхолин и называются холинергическими. Некоторые нейроны продолговатого мозга и варолиева моста синтезируют норадреналин и их относят к норадренергическим. В продолговатом мозге имеются также серотонинергические нейроны, секретирующие серотонин, в мозжечке – ГАМК-ергические, которые выделяют гамма-аминомасляную кислоту (ГАМК). В среднем мозге находятся дофаминергические нейроны, выделяющие дофамин.
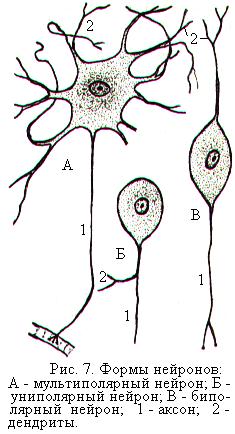
Классификация нейронов по их внешней структуре основана на форме клеток и количестве отходящих от них отростков. По форме различают нейроны веретеновидные, звёздчатые, пирамидные и т.д. По количеству отходящих отростков нервные клетки подразделяют на униполярные, т.е. с одним отростком, биполярные с двумя отростками и мультиполярные со многими отростками (рис. 7), к которым относятся большинство нейронов ЦНС.
Биохимия нервной клетки. Аксонный транспорт
Оболочка ядра нервной клетки имеет большое количество впячиваний и выпячиваний, во много раз увеличивающих её площадь. Она также пронизана множеством пор, через которые транспортируется РНК к рибосомам, находящимся в цитоплазме нейрона. Рибосомы, связанные с мембраной гранулярной эндоплазматической сети, обеспечивают мощную белоксинтезирующую функцию нервных клеток. На фотографиях, полученных с электронного микроскопа, видно, что при функциональных нагрузках или при заболеваниях увеличивается количество свободных рибосом и уменьшается число рибосом, связанных с эндоплазматической сетью.
Синтезируемый в нейроне белок идёт на поддержание и обновление многочисленных нервных отростков. Белки тела нейрона разделяют на два типа: быстро обновляющиеся (с короткой продолжительностью жизни) и медленно обновляющиеся. Быстро обновляющиеся белки живут около 1 суток, а медленно обновляющиеся – до 15 суток. Медленно обновляющиеся белки составляют основную часть структуры тела клетки, а быстро обновляющиеся поступают из тела нейрона в его отростки. Кроме строительных белков в нервной клетке синтезируются белки, обеспечивающие передачу нервных импульсов, а также белки, необходимые для нормального функционирования иннервируемой ткани, т.е. ткани, в которой заканчиваются отростки нейрона.
Нейроны гипоталамуса способны к нейросекреции, т.е. могут синтезировать биологически активные вещества – нейрогормоны. Нейрогормоны с током крови или по аксонам нейронов гипоталамуса поступают в гипофиз, влияют на выделение гормонов гипофизом и тем самым участвуют в регуляции функций организма.
Под электронным микроскопом в нервной клетке видно большое количество различных пузырьков. В этих пузырьках содержатся многие вещества, синтезированные в нейроне. Разные по своей химической природе вещества, упакованные в пузырьки, транспортируются по нервным волокнам. В нервных окончаниях в пузырьках содержится медиатор – химическое вещество, с помощью которого нервный импульс передаётся с нейрона на нейрон.
В отростках нервной клетки имеются особые нити – нейрофибриллы. Они состоят из микротрубочек и нейрофиламентов, представляющих собой длинные волокнистые структуры белковой природы. Система нейрофибрилл принимает непосредственное участие в обеспечении транспорта веществ в нервных отростках. Эти перемещения веществ хорошо изучены в аксонах и получили название аксонный транспорт. Существует два вида аксонного транспорта: быстрый и медленный.
Быстрый аксонный транспорт – это перемещение пузырьков, митохондрий и некоторых белков от тела клетки по аксону к его окончанию или в обратном направлении со скоростью 250-400 мм в сутки. Этот транспорт не нарушается при отделении аксона от тела нейрона, но прекращается при разрушении внутриаксонных микротрубочек и нейрофиламентов. Быстрый аксонный транспорт пузырьков от тела нейрона к окончанию аксона называется антероградным транспортом. Быстрый аксонный транспорт в обратном направлении, т.е. от окончания аксона к телу нейрона, называют ретроградным транспортом.
Медленный аксонный транспорт – это перемещение всей массы белков от тела нейрона к окончанию аксона со скоростью 1-4 мм в сутки. Этот транспорт не нарушается при разрушении микротрубочек, но прекращается при отделении аксона от тела нейрона. Он имеет большое зна-
чение в процессах роста нервных отростков и их восстановления после повреждений.
Нейроглия. Гематоэнцефалический барьер
Нейроглию подразделяют на макроглию и микроглию. Клетки макроглии – астроциты, олигодендроциты и эпендимоциты выполняют в нервной системе важные функции. Астроциты осуществляют опорную функцию, заполняя пространство между нейронами, а также замещая погибшие нервные клетки. На нейроне обычно оканчиваются аксоны многих других нервных клеток, и все они изолированы друг от друга астроцитами. Астроциты очень часто заканчиваются своими отростками на кровеносных сосудах, образуя так называемые сосудистые ножки. Астроциты также способны уничтожать микробы и вредные вещества.
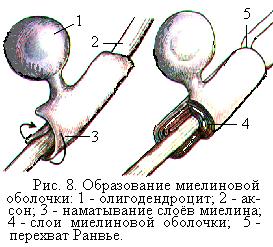 Олигодендроциты образуют мякотные (миелиновые) оболочки вокруг нервных волокон (рис. 8). Олигодендроциты также окружают со всех сторон нейроны и обеспечивают для них питание и выделение.
Олигодендроциты образуют мякотные (миелиновые) оболочки вокруг нервных волокон (рис. 8). Олигодендроциты также окружают со всех сторон нейроны и обеспечивают для них питание и выделение.
Эпендимоциты – это эпителиальные клетки, выстилающие полости желудочков мозга. Один отросток эпендимоцита доходит до кровеносного сосуда. Полагают, что эпендимоциты являются посредниками между кровеносным сосудом и полостью мозговых желудочков, заполненных спинномозговой жидкостью.
Источником клеток микроглии служат мозговая оболочка, стенка кровеносных сосудов и сосудистая оболочка желудочков мозга. Клетки микроглии способны передвигаться. Они осуществляют захват и последующую переработку попавших в организм микробов, инородных веществ, а также отмерших элементов мозга. Скопления клеток микроглии часто наблюдаются около участков повреждённого мозгового вещества.
Большую роль клетки нейроглии играют в осуществлении барьера между кровью и мозгом, так называемого гематоэнцефалического барьера. Не все вещества, попадающие в кровь, могут проникнуть в мозг. Они задерживаются гематоэнцефалическим барьером, который предохраняет мозг от поступления из крови различных вредных для него веществ, а также многих бактерий. В выполнении барьерных функций наряду с другими структурными образованиями участвуют астроциты. Сосудистые ножки астроцитов со всех сторон окружают кровеносный капилляр, плотно соединяясь между собой.
Если по каким-то причинам гематоэнцефалический барьер нарушается, то микробы или ненужные вещества могут проникнуть в мозг и в первую очередь в цереброспинальную жидкость. Цереброспинальная, или спинномозговая жидкость, или ликвор – это внутренняя среда мозга, поддерживающая его солевой состав, участвующая в питании мозговых клеток и удалении из них продуктов распада. Она также поддерживает внутричерепное давление, является гидравлической подушкой мозга, предохраняющей нервные клетки от повреждений при ходьбе, беге, прыжках и других движениях.
Цереброспинальная жидкость заполняет желудочки головного мозга, центральный канал спинного мозга, пространства между оболочками как головного, так и спинного мозга. Она постоянно циркулирует. Нарушение её циркуляции ведёт к расстройствам деятельности ЦНС. Количество цереброспинальной жидкости у взрослого человека равно 120-150 мл. В ней отсутствуют ферменты и иммунные тела, содержится небольшое количество лимфоцитов. В ней меньше, чем в крови, белков и примерно такое же, как в крови, содержание минеральных солей.
Многие вещества, находящиеся в крови или искусственно вводимые в кровь, совсем не попадают в спинномозговую жидкость и соответственно в клетки мозга. Поэтому определённые лекарства, например некоторые антибиотики, для лечения нейронов спинного или головного мозга приходится вводить непосредственно в цереброспинальную жидкость, прокалывая оболочки спинного мозга. Вместе с тем, такие вещества как алкоголь, хлороформ, морфий, столбнячный токсин легко проникают через гематоэнцефалический барьер в цереброспинальную жидкость и быстро действуют на нейроны мозга. Проницаемость гематоэнцефалического барьера регулируется центральной нервной системой. Благодаря этому мозг может в определённой мере сам регулировать собственное функциональное состояние.
МЕМБРАННЫЕ ПОТЕНЦИАЛЫ НЕРВНЫХ КЛЕТОК
Плазматическая мембрана нейрона
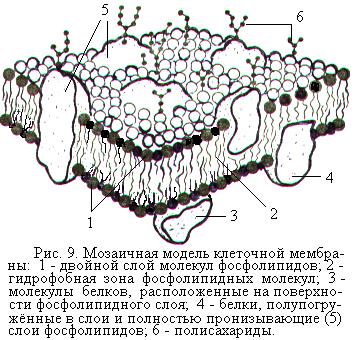
Плазматической, или цитоплазматической, или клеточной мембраной, или плазмалеммой называют мембрану, отделяющую цитоплазму клетки от наружной среды. Основу плазматической мембраны нейрона составляет двойной слой молекул фосфолипидов (рис. 9). Главная функция фосфолипидов состоит в поддержании механической стабильности плазмалеммы и придании ей водоотталкивающих свойств. В бимолекулярный слой фосфолипидов погружены частично или пронизывают его насквозь молекулы белков. Выступающие из двойного слоя фосфолипидов части молекул белков образуют третий слой плазматической мембраны нейрона. Белки, пронизывающие насквозь липидную основу мембраны, образуют каналы для воды и ионов, формируют ионные насосы.
Многие белки мембраны представляют собой гликопротеиды с полисахаридными цепочками, которые выступают над наружной поверхно-
стью. Вместе с другими углеводными молекулами эти цепочки образуют тонкий слой на поверхности клетки, называемый гликокаликсом. Липидные и белковые компоненты находятся в мембране в жидком состоянии и поэтому обладают значительной подвижностью (текучестью). Такая модель строения плазмалеммы получила название жидкой мозаичной мембраны.
 Основные функции плазматической мембраны – барьерная, транспортная, регуляторная и каталитическая. Барьерная функция заключается в ограничении диффузии через мембрану растворимых в воде соединений. Это необходимо для защиты нейрона от чужеродных, вредных веществ и сохранения внутри клетки определённой концентрации нужных веществ. Регуляция плазматической мембраной обмена веществ между цитоплазмой и окружающей клетку средой особенно важна для нейронов по следующим причинам. Во-первых, мембрана регулирует транспорт веществ, которые непосредственно связаны с нервной сигнализацией. Во-вторых, мембрана служит местом возникновения электрических потенциалов, лежащих в основе нервных импульсов. В-третьих, она служит местом действия пептидов и гормонов. И, наконец, участки плазмалеммы образуют синапсы, где сигналы передаются от одной клетки к другой. Сложный ферментный комплекс мембраны обеспечивает восприятие информации из внеклеточной среды и передачу её внутрь клетки.
Основные функции плазматической мембраны – барьерная, транспортная, регуляторная и каталитическая. Барьерная функция заключается в ограничении диффузии через мембрану растворимых в воде соединений. Это необходимо для защиты нейрона от чужеродных, вредных веществ и сохранения внутри клетки определённой концентрации нужных веществ. Регуляция плазматической мембраной обмена веществ между цитоплазмой и окружающей клетку средой особенно важна для нейронов по следующим причинам. Во-первых, мембрана регулирует транспорт веществ, которые непосредственно связаны с нервной сигнализацией. Во-вторых, мембрана служит местом возникновения электрических потенциалов, лежащих в основе нервных импульсов. В-третьих, она служит местом действия пептидов и гормонов. И, наконец, участки плазмалеммы образуют синапсы, где сигналы передаются от одной клетки к другой. Сложный ферментный комплекс мембраны обеспечивает восприятие информации из внеклеточной среды и передачу её внутрь клетки.
Плазмалемма нейрона легко проницаема для жирорастворимых веществ, молекулы которых проникают через двойной слой фосфолипидов. Крупные водорастворимые молекулы, а также анионы органических кислот, практически совсем не проходят через мембрану. В то же время в мембране нервного волокна существуют каналы, проницаемые для воды, для малых молекул водорастворимых веществ и для малых ионов.
Ионные каналы. Транспорт ионов
Каналы, проницаемые для ионов, или ионные каналы, образованы молекулами белков, пронизывающими бимолекулярный слой фосфолипидов. Ионные каналы мембраны обладают двумя важнейшими свойствами: 1) избирательно пропускают одни ионы и не пропускают другие; 2) обладают электровозбудимостью, т.е. способностью открываться и закрываться в ответ на изменения электрического заряда на мембране. Избирательность ионных каналов определяется геометрическими размерами иона и химической природой белка, образующего стенки канала.
Особое значение имеют каналы, проницаемые для ионов натрия, калия, хлора и кальция. В мембране нейрона имеются специфические селективные натриевые, калиевые, хлорные и кальциевые каналы, т.е. каналы, избирательно пропускающие только ионы Nа+ или K+ или Cl - или Са2+. Эти каналы имеют механизмы для их открывания и закрывания. От состояния (открытого или закрытого) определённых ионных каналов мембраны зависит величина электрического заряда (потенциала) на мембране клетки. Кроме специфических селективных, в мембране находятся неспецифические каналы для ионной утечки, каждый из которых проницаем больше всего для ионов калия и в меньшей степени для ионов натрия и хлора. Эти каналы не имеют механизмов для их закрытия и открывания и почти не меняют своего состояния при электрических воздействиях на мембрану нейрона.
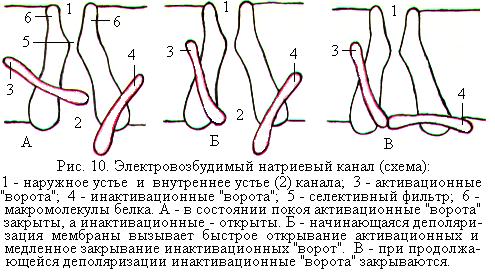 |
Принято считать, что ионный канал имеет наружное и внутреннее расширение, так называемые «устья», и короткий суженный участок, или селективный фильтр (рис. 10). Через селективный фильтр проходят только те ионы, которые соответствуют по размерам и свойствам. Ионные каналы обладают воротными механизмами, или «воротами», с помощью которых они могут быть открытыми или закрытыми. «Ворота» расположены в области внутреннего конца канала. Они могут находиться только в двух положениях: или полностью открыты или полностью закрыты. Поэтому ионная проницаемость одиночного открытого канала является постоянной величиной. Суммарная проводимость мембраны для того или иного иона определяется числом одновременно открытых каналов, проницаемых для данного иона.
Через открытые каналы ионы могут проходить через клеточную мембрану пассивным и активным способом. На пассивный транспорт ве-
ществ клетка своей энергии не затрачивает. Пассивный транспорт веществ осуществляется по концентрационному и электрическому градиентам путём фильтрации, диффузии и осмоса. Активный транспорт ионов через мембрану клетки идёт по ионным каналам, требует затраты энергии клеткой и осуществляется с помощью так называемых ионных насосов. Через один ионный канал может проходить от 10 млн. до 100 млн. ионов в секунду.
Перенос ионов против градиентов их концентраций, т.е. из раствора с меньшей концентрацией в раствор с большей концентрацией этих ионов, называется активным ионным транспортом в отличие от пассивного транспорта, часто называемого утечкой ионов. В мембране клетки имеется натрий-калиевый насос, который способен против градиента концентрации откачивать ионы натрия из цитоплазмы во внеклеточную среду в обмен на ионы калия. Натрий-калиевый насос работает, потребляя энергию АТФ. Молекулярный механизм насоса функционирует лишь в случае присоединения к нему снаружи клетки ионов калия, а изнутри клетки – ионов натрия. В мембране клетки имеются насосы и для других ионов, например, для ионов кальция, хлора.
Таким образом, в живой клетке есть две системы движения ионов через мембрану. Одна из них – пассивный ионный транспорт – осуществляется по градиенту концентрации ионов и не требует затраты энергии клеткой. Благодаря ему возникает потенциал покоя и потенциал действия. Он может привести в конечном итоге к выравниванию концентрации ионов натрия и калия по обе стороны клеточной мембраны. Однако в живой клетке этого не происходит вследствие наличия второй системы движения ионов – активного ионного транспорта.
Активный транспорт – это результат работы натрий-калиевого насоса, благодаря которому поддерживается постоянство разности концентраций ионов между цитоплазмой и внеклеточной жидкостью в покое, а также восстанавливается исходная разность ионных концентраций, нарушающаяся при каждом возбуждении нейрона. Активный ионный транспорт возможен лишь при условии затраты клеткой энергии АТФ, пополняемой в результате процессов обмена веществ.
Мембранный потенциал покоя
Любая живая клетка покрыта полупроницаемой мембраной, через которую осуществляется пассивное движение и активный избирательный транспорт положительно и отрицательно заряженных ионов. Благодаря этому переносу между наружной и внутренней поверхностью мембраны имеется разность электрических зарядов (потенциалов) – мембранный потенциал. Существует три отличающихся друг от друга проявления мембранного потенциала – мембранный потенциал покоя, местный потенциал, или локальный ответ, и потенциал действия.
Если на клетку не действуют внешние раздражители, то мембранный потенциал долго сохраняется постоянным. Мембранный потенциал такой покоящейся клетки называется мембранным потенциалом покоя. Для наружной поверхности мембраны клетки потенциал покоя всегда положителен, а для внутренней поверхности клеточной мембраны всегда отрицателен. Принято измерять потенциал покоя на внутренней поверхности мембраны, т.к. ионный состав цитоплазмы клетки более стабилен, чем межклеточной жидкости. Величина потенциала покоя относительно постоянна для каждого типа клеток. Для поперечнополосатых мышечных клеток она составляет от –50 до – 100 мВ, а для нервных клеток от –60 до – 70 мВ.
Причиной возникновения потенциала покоя является разная концентрация катионов и анионов снаружи и внутри клетки, а также избирательная проницаемость для них клеточной мембраны. Цитоплазма покоящейся нервной и мышечной клетки содержит примерно в 20-100 раз больше катионов калия, в 5-15 раз меньше катионов натрия и в 20-100 раз меньше анионов хлора, чем внеклеточная жидкость.
В состоянии покоя практически все натриевые каналы мембраны клетки закрыты, а большинство калиевых каналов открыто. Всякий раз, когда ионы калия наталкиваются на открытый канал, они проходят через мембрану. Поскольку внутри клетки ионов калия гораздо больше, то осмотическая сила выталкивает их из клетки. Вышедшие катионы калия увеличивают положительный заряд на наружной поверхности клеточной мембраны. В результате выхода ионов калия из клетки должна была бы вскоре уравняться их концентрация внутри и вне клетки. Однако этому препятствует электрическая сила отталкивания положительных ионов калия от положительно заряженной наружной поверхности мембраны.
Чем больше становится величина положительного заряда на наружной поверхности мембраны, тем труднее ионам калия проходить из цитоплазмы через мембрану. Ионы калия будут выходить из клетки до тех пор, пока сила электрического отталкивания не станет равной силе осмотического давления К+. При таком уровне потенциала на мембране вход и выход ионов калия из клетки находятся в равновесии, поэтому электрический заряд на мембране в этот момент называется калиевым равновесным потенциалом. Для нейронов он равен от –80 до –90 мВ.
Поскольку в покоящейся клетке почти все натриевые каналы мембраны закрыты, то ионы Nа+ поступают в клетку по концентрационному градиенту в незначительном количестве. Они лишь в очень малой степени возмещают потерю положительного заряда внутренней средой клетки, вызванную выходом ионов калия, но не могут эту потерю существенно компенсировать. Поэтому проникновение в клетку (утечка) ионов натрия приводит лишь к незначительному снижению мембранного потенциала, вследствие чего мембранный потенциал покоя имеет несколько меньшую величину по сравнению с калиевым равновесным потенциалом.
|
|
|
|
|
Дата добавления: 2014-12-26; Просмотров: 855; Нарушение авторских прав?; Мы поможем в написании вашей работы!