КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Строение синапса. Синаптическая передача возбуждения ….….. 23 4 страница
|
|
|
|
У ребёнка в первые 3 месяца внутриутробной жизни спинной мозг занимает всю длину позвоночного канала. Затем, вследствие более быстрого роста позвоночника, спинной мозг отстаёт в росте и его нижние отделы – поясничный, крестцовый и копчиковый располагаются в более высоко расположенных отделах позвоночника.
Вдоль спинного мозга проходят передняя срединная щель и задняя срединная борозда, которые делят его на две симметричные половины. На боковых поверхностях спинного мозга симметрично выходят передние корешки и входят задние корешки спинномозговых нервов (рис. 28). Участок спинного мозга, который соответствует каждой паре корешков, называется сегментом. Сегменты обозначаются латинскими буквами в соответствии с отделом спинного мозга: шейный (cervicalis) – С, грудной (thoracalis) – Th, поясничный (lumbalis) – L, крестцовый (sacralis) – S, копчиковый (cоccygeus) – Со. Рядом с буквой, которой называется отдел мозга, ставят цифру, обозначающую номер сегмента в данном
отделе. Например, L2 – второй поясничный сегмент. Всего в спинном мозге человека – 31 сегмент.
Шейный отдел спинного мозга имеет 8 сегментов и переходит на уровне 7-го шейного позвонка в грудной отдел, состоящий из 12 сегментов. Нижняя граница грудного отдела лежит на уровне 10-11-го грудного позвонка. Поясничный отдел состоит из 5 сегментов, нижняя граница которых проходит на уровне 12-го грудного позвонка. Крестцовый отдел (5 сегментов) на уровне 1-го поясничного позвонка переходит в копчиковый отдел, чаще всего состоящий из 1 сегмента. Копчиковый отдел заканчивается на уровне нижнего края 1-го поясничного позвонка.
Место выхода спинномозговых корешков из мозга не соответствует уровню межпозвоночных отверстий. Поэтому корешки, прежде чем выйти из позвоночного канала, направляются вниз вдоль спинного мозга и в поясничном отделе позвоночника образуют пучок – конский хвост.
Задние корешки состоят из аксонов чувствительных нейронов, тела которых находятся в спинномозговых нервных узлах, или ганглиях, расположенных по обе стороны у входа или во входе в позвоночник - межпозвоночном отверстии. Чувствительные нейроны имеют один отросток, который сразу после отхода от тела клетки делится на две ветви. Одна из них проводит нервные импульсы от рецепторов к телу клетки и является дендритом, другая – от тела нервной клетки к нейронам спинного мозга и является аксоном. Проведение нервных импульсов из одной ветви в другую может проходить и без участия тела чувствительного нейрона.
Передние корешки образованы аксонами двигательных нейронов, или мотонейронов, тела которых расположены в передних рогах спинного мозга. В грудном отделе в передние корешки кроме аксонов двигательных нейронов входят аксоны вегетативных нейронов, тела которых находятся в боковых рогах спинного мозга. Передние и задние корешки (спинномозговые корешки) в области межпозвоночного отверстия соединяются в спинномозговой нерв. У человека 31 пара спинномозговых нервов, соответствующих 31 сегменту спинного мозга.
В опытах на животных с перерезкой отдельных корешков было установлено, что каждый сегмент спинного мозга иннервирует три поперечных отрезка, или метамера, тела: свой собственный, соседний сверху и соседний снизу. Это создаёт надёжность иннервации, т.к. для того чтобы лишить чувствительности участок тела, необходимо перерезать три смежных задних корешка. Точно также сокращение мышц и движение участка тела исчезает при повреждении или перерезке трёх соседних передних корешков.
На поперечном разрезе спинного мозга видно, что его очень узкий центральный канал окружён серым веществом (рис. 28). Оно образовано телами и дендритами нейронов и напоминает форму летящей бабочки. Выступающие части серого вещества называют передними и задними ро гами. 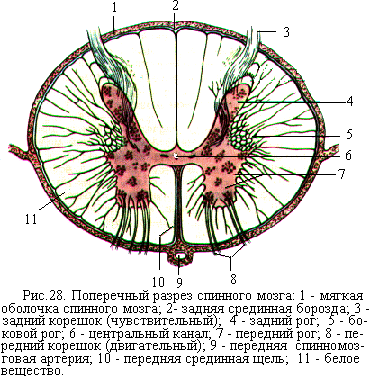 В грудном отделе и верхней части поясничного отдела между передними и задними рогами есть ещё и боковые рога.
В грудном отделе и верхней части поясничного отдела между передними и задними рогами есть ещё и боковые рога.
В спинном мозге человека содержится примерно 13 млн. нейронов. Из них только около 3 % составляют эфферентные клетки – двигательные, или мотонейроны, и вегетативные нейроны. Остальные 97 % приходятся на долю вставочных нейронов, или интернейронов. Мотонейроны спинного мозга подразделяются на два основных типа: альфа-мотонейроны и гамма-мотонейроны.
Серое вещество спинного мозга со всех сторон окружено белым веществом. Белое вещество делится на три парных канатика – передний, боковой и задний. Передний канатик расположен между передней срединной щелью и передним рогом серого вещества, боковой канатик – между передним и задним рогом, задний канатик – между задним рогом и задней срединной бороздой. Белое вещество спинного мозга образовано в основном продольно идущими нервными волокнами, объединёнными в пучки, которые являются проводящими путями для нервных импульсов.
Функции спинного мозга
Спинной мозг у человека является низшим отделом ЦНС, имеющим сегментарную организацию. Сегментарность заключается в том, что: 1) сегменты спинного мозга иннервируют участки тела, расположенные на том же уровне (шейные – шею, грудные – грудь и т.д.); 2) нейроны спинного мозга напрямую связаны с рабочими органами в отличие от нейронов надсегментарных центров, например коры больших полушарий, которые управляют деятельностью мышц через сегментарные центры, в том числе и через спинной мозг. Спинной мозг у человека имеет значительно меньшую самостоятельность по сравнению со спинным мозгом животных.
Спинной мозг выполняет интегративную, проводниковую и рефлекторную функции. Интегративная функция заключается в объединении (интеграции) поступающей информации от рецепторов. Она у спинного мозга проявляется слабо. Интегративная функция более значительна у
вышележащих отделов ЦНС и наиболее выражена у коры больших полушарий.
Проводниковая функция у спинного мозга выражена очень хорошо и осуществляется благодаря проводящим путям. Различают восходящие и нисходящие проводящие пути. По восходящим путям импульсы поступают от нижележащих сегментов спинного мозга к вышележащим сегментам и к головному мозгу, а по нисходящим путям – в обратном направлении.
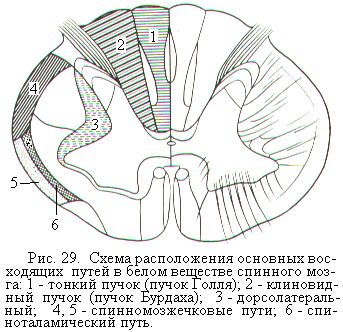 Основные восходящие проводящие пути спинного мозга идут в составе задних и боковых канатиков (рис. 29). Их называют чувствительными путями, т.к. они проводят импульсы от температурных, болевых, тактильных рецепторов, от рецепторов кожи, от проприорецепторов мышц, сухожилий и связок, от рецепторов, сигнализирующих о положении тела, пассивных движениях тела и вибрации. Наиболее важными восходящими путями являются расположенные в задних канатиках спинного мозга тонкий пучок (пучок Голля) и клиновидный пучок (пучок Бурдаха), ведущие в продолговатый мозг. В боковых канатиках расположены спинно-мозжечковые пути, ведущие в мозжечок и спиноталамический путь, ведущий в ядра таламуса.
Основные восходящие проводящие пути спинного мозга идут в составе задних и боковых канатиков (рис. 29). Их называют чувствительными путями, т.к. они проводят импульсы от температурных, болевых, тактильных рецепторов, от рецепторов кожи, от проприорецепторов мышц, сухожилий и связок, от рецепторов, сигнализирующих о положении тела, пассивных движениях тела и вибрации. Наиболее важными восходящими путями являются расположенные в задних канатиках спинного мозга тонкий пучок (пучок Голля) и клиновидный пучок (пучок Бурдаха), ведущие в продолговатый мозг. В боковых канатиках расположены спинно-мозжечковые пути, ведущие в мозжечок и спиноталамический путь, ведущий в ядра таламуса.
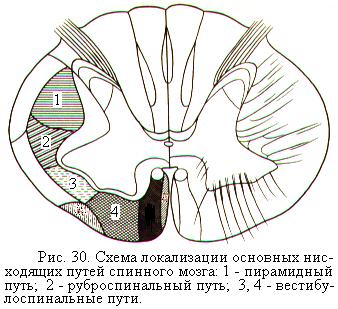
Основные нисходящие проводящие пути спинного мозга идут в составе передних и боковых канатиков. Они получили название двигательных путей, т.к. проводят к мышцам импульсы, которые обеспечивают произвольные движения, тонус скелетных мышц, поддержание позы и равновесия тела. Наиболее важными нисходящими путями являются пирамидные и экстрапирамидные пути (рис. 30). По пирамидным путям поступают импульсы от двигательной зоны коры больших полушарий к мотонейронам спинного мозга. В состав пирамидных путей подключаются также волокна вегетативной нервной системы, несущие эфферентные импульсы к внутренним органам.
От пирамидных путей отходят ответвления к подкорковым ядрам, ядрам ствола мозга и к ретикулярной формации. От этих ядер и ретикулярной формации отходят экстрапирамидные пути. Они передают влияния, регулирующие мышечный тонус при выполнении произвольных движений. Важнейшими из экстрапирамидных путей являются руброспинальный тракт и вестибулоспинальные тракты (рис. 30).
 Рефлекторная функция спинного мозга осуществляется благодаря передаче нервных импульсов с афферентных путей на эфферентные, в результате чего происходит ответная реакция на раздражение. Связь спинного мозга с рецепторами и рабочими органами осуществляется посредством нервных волокон, входящих в спинной мозг и выходящих из него в составе спинномозговых корешков. Распределение функций этих волокон подчиняется закону Белла-Мажанди, а именно: все чувствительные (афферентные) волокна входят в спинной мозг через задние корешки, а все двигательные (эфферентные) волокна выходят через передние корешки. В задних корешках волокон примерно в 5 раз больше, чем в передних.
Рефлекторная функция спинного мозга осуществляется благодаря передаче нервных импульсов с афферентных путей на эфферентные, в результате чего происходит ответная реакция на раздражение. Связь спинного мозга с рецепторами и рабочими органами осуществляется посредством нервных волокон, входящих в спинной мозг и выходящих из него в составе спинномозговых корешков. Распределение функций этих волокон подчиняется закону Белла-Мажанди, а именно: все чувствительные (афферентные) волокна входят в спинной мозг через задние корешки, а все двигательные (эфферентные) волокна выходят через передние корешки. В задних корешках волокон примерно в 5 раз больше, чем в передних.
Спинной мозг выполняет много рефлекторных функций. В нём находятся центры, осуществляющие движения всех скелетных мышц за исключением мышц, расположенных на голове. Также в спинном мозге расположены центры всех рефлексов мочеполовой системы и прямой кишки, терморегуляторных рефлексов, центры, регулирующие обмен веществ, сокращение диафрагмы, центры большинства сосудистых рефлексов.
Степень проявления рефлексов зависит от наличия связи структур спинного мозга со структурами головного мозга. После спинализации, т.е. отделения спинного мозга от головного у животных посредством перерезки или вследствие травмы у человека, исчезают многие сложные формы активности, создаваемые спинным мозгом - спинальный шок. Продолжительность спинального шока у человека от нескольких недель до 4-5 месяцев. Когда шок проходит, рефлексы восстанавливаются.
Рефлексы, протекающие с участием спинного мозга, подразделяются на двигательные и вегетативные. Двигательные рефлексы осуществляются с помощью альфа-мотонейронов передних рогов. Альфа-мото-нейроны – это крупные клетки, от которых отходят толстые быстропроводящие нервные волокна. Они осуществляют передачу к скелетным мышцам импульсов, выработанных в спинном мозге. Аксон каждого мотонейрона, вступая в мышцу, многократно делится, и может охватывать своими окончаниями до нескольких сотен экстрафузальных мышечных волокон, образуя с ними двигательную, или нейромоторную единицу.
Импульсы, поступающие от альфа-мотонейронов, вызывают сокращение скелетных мышц.
Потенциалы действия в альфа-мотонейроне возникают с частотой, не превышающей 200-300 имп/с, и со следовой гиперполяризацией длительностью от 50 до 150 мс. В зависимости от частоты воспроизведения импульсов и длительности следовой гиперполяризации альфа-мото-нейроны разделяют на фазические (фазные) и тонические. Фазическими мотонейронами иннервируются более быстрые, «белые» мышцы, тоническими – более медленные «красные».
Второй тип мотонейронов спинного мозга – гамма-мотонейроны посылают тонкие волокна к интрафузальным (внутриверетённым) мышечным волокнам, которые являются мышечными рецепторами. Импульсы, поступающие от гамма-мотонейронов, не вызывают сокращение скелетных мышц, а изменяют чувствительность мышечных рецепторов.
В спинном мозге замыкается огромное количество рефлекторных дуг, с помощью которых регулируются функции организма. Типичные спинномозговые (спинальные) рефлексы, т.е. рефлексы, присущие самому спинному мозгу, можно изучить в чистом виде только после отделения спинного мозга от головного у животного (спинальное животное) или у человека, у которого в результате травмы произошёл разрыв спинного мозга на границе с продолговатым мозгом. Первым следствием нарушения целостности спинного мозга является спинальный шок. Он проявляется в резком падении возбудимости и в угнетении рефлекторных функций всех рефлекторных центров, расположенных ниже места перерезки или разрыва.
Спустя несколько недель, а то и месяцев у человека спинальный шок медленно проходит и постепенно восстанавливается рефлекторная деятельность скелетных мышц, величина кровяного давления, рефлексы мочеиспускания, дефекации и ряд половых рефлексов. Это свидетельствует о том, что центры этих рефлексов располагаются в спинном мозге.
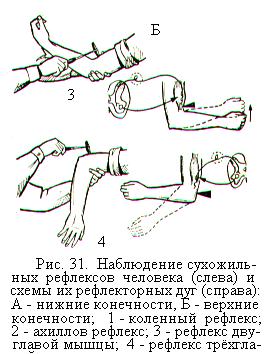
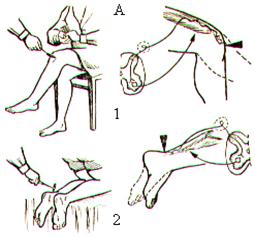
Первыми после отделения головного мозга от спинного и снятия шока появляются сгибательные и разгибательные рефлексы. Вслед за ними обнаруживаются сухожильные рефлексы (рис. 31), дуги которых являются моносинаптическими, т.е. имеющими только один синапс. Моносинаптическая дуга состоит из чувствительного нейрона, дендриты которого идут от рецепторов растяжения – мышечных веретён, и эфферентного нейрона, оканчивающегося на мышечных волокнах. В спинном мозге меньше моносинаптических дуг и больше полисинаптических.
Сухожильные рефлексы легко вызываются с помощью короткого удара по сухожилию и имеют важное диагностическое значение в неврологической практике. Ответная рефлекторная реакция на удар по сухожилию проявляется в виде резкого сокращения мышцы. Особенно выражены сухожильные рефлексы в мышцах разгибателях ноги (рис. 31, А), таких как четырёхглавая мышца бедра (коленный рефлекс) или трёхглавая мышца голени (ахиллов рефлекс). Сухожильные рефлексы можно вызвать и в мышцах-сгибателях. На руке они чётко проявляются на двуглавой и трёхглавой мышцах (рис.31, Б).
|
 Важное значение для поддержания выпрямленной позы и стояния человека имеют рефлексы растяжения. Под влиянием силы тяжести раздражаются рецепторы растяжения (мышечные веретёна) в мышце. Так, при стоянии четырёхглавая мышца бедра подвергается растяжению из-за тенденции колена сгибаться под влиянием гравитации. От мышечных веретён четырёхглавой мышцы поступают длительное время асинхронные импульсы к альфа-мотонейронам спинного мозга. В ответ альфа-мотонейроны посылают тоже асинхронные импульсы к экстрафузальным волокнам четырёхглавой мышцы. В результате этого мышца отвечает плавным длительным сокращением, автоматически противодействующим силе тяжести. Рефлексы растяжения иногда относят к группе тонических или позных рефлексов, т.к. их объединяет длительное поддержание рефлекторного сокращения разгибательных и реже сгибательных мышц без заметного их утомления. Тонические рефлексы направлены на поддержание определённой позы. В их осуществлении кроме механизмов спинного мозга участвуют двигательные центры головного мозга.
Важное значение для поддержания выпрямленной позы и стояния человека имеют рефлексы растяжения. Под влиянием силы тяжести раздражаются рецепторы растяжения (мышечные веретёна) в мышце. Так, при стоянии четырёхглавая мышца бедра подвергается растяжению из-за тенденции колена сгибаться под влиянием гравитации. От мышечных веретён четырёхглавой мышцы поступают длительное время асинхронные импульсы к альфа-мотонейронам спинного мозга. В ответ альфа-мотонейроны посылают тоже асинхронные импульсы к экстрафузальным волокнам четырёхглавой мышцы. В результате этого мышца отвечает плавным длительным сокращением, автоматически противодействующим силе тяжести. Рефлексы растяжения иногда относят к группе тонических или позных рефлексов, т.к. их объединяет длительное поддержание рефлекторного сокращения разгибательных и реже сгибательных мышц без заметного их утомления. Тонические рефлексы направлены на поддержание определённой позы. В их осуществлении кроме механизмов спинного мозга участвуют двигательные центры головного мозга.
Ритмические рефлексы, или рефлексы мышц-антагонистов, лежат в основе правильных попеременных движений при ходьбе, беге, чесании и т.д. Эти рефлексы характеризуются тем, что при возбуждении мотонейронов мышц-сгибателей одновременно происходит торможение мотонейронов мышц-разгибателей. При этом в конечности другой стороны наблюдаются обратные явления. Центры рефлекса ходьбы лежат в поясничных сегментах спинного мозга.
К двигательным рефлексам спинного мозга относят также висцеромоторные рефлексы. Они возникают при определённых раздражениях внутренних органов и характеризуются появлением двигательных реак-
ций мышц грудной и брюшной стенки, мышц-разгибателей спины (например, при икоте).
Двигательные рефлексы спинного мозга называют также соматическими, т.к. они проявляются в деятельности скелетных мышц. Кроме их осуществления, спинной мозг играет важную роль в рефлекторной регуляции внутренних органов, являясь центром многих вегетативных рефлексов. Вегетативные рефлексы спинного мозга разделяют на симпатические и парасимпатические. В боковых рогах сегментов грудного отдела и верхних сегментов поясничного отдела спинного мозга расположены центры симпатического отдела, а в крестцовом отделе – центры парасимпатического отдела вегетативной нервной системы. Эти центры участвуют в регуляции пищеварения, дыхания, кровообращения, мочеиспускания, дефекации, осуществляют некоторые половые рефлексы.
ПРОДОЛГОВАТЫЙ МОЗГ И ВАРОЛИЕВ МОСТ
Строение продолговатого мозга и варолиева моста
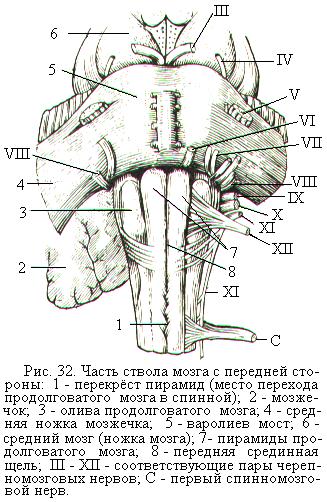 Продолговатый мозг является продолжением спинного и в нижней части похож на него по строению и форме (рис. 32). В продолговатом мозге частично сохраняется сегментарное строение, типичное для спинного мозга. Вверху продолговатый мозг переходит в варолиев мост, а боковые его отделы переходят в нижние ножки мозжечка, который находится сзади. В продолговатом мозге продолжаются, переходя со спинного мозга, передняя срединная щель, задняя срединная борозда, а внутри центральный канал. По бокам передней срединной щели располагаются пирамиды, образованные пучками проводящих путей. Сбоку от пирамид с каждой стороны продолговатого мозга находятся оливы.
Продолговатый мозг является продолжением спинного и в нижней части похож на него по строению и форме (рис. 32). В продолговатом мозге частично сохраняется сегментарное строение, типичное для спинного мозга. Вверху продолговатый мозг переходит в варолиев мост, а боковые его отделы переходят в нижние ножки мозжечка, который находится сзади. В продолговатом мозге продолжаются, переходя со спинного мозга, передняя срединная щель, задняя срединная борозда, а внутри центральный канал. По бокам передней срединной щели располагаются пирамиды, образованные пучками проводящих путей. Сбоку от пирамид с каждой стороны продолговатого мозга находятся оливы.
Задняя поверхность продолговатого мозга разделена задней срединной бороздой. По бокам от неё расположены тонкий и клиновидный пучки, образующие задний канатик. Задние канатики продолговатого мозга являются продолжением задних канатиков спинного мозга. Вверху задние канатики переходят в нижние ножки мозжечка.
Продолговатый мозг участвует в образовании одной из полостей головного мозга – IV-го желудочка. Продолговатый мозг и мост образуют дно IV-го желудочка – ромбовидную ямку (рис. 33). Внизу IV желудочек сообщается с центральным каналом спинного мозга, вверху переходит в водопровод среднего мозга.
Варолиев мост расположен над продолговатым мозгом в виде поперечного белого вала (рис. 32). Сверху он переходит в ножки мозга, а боковые его отделы переходят в средние ножки мозжечка. Мост и мозжечок часто объединяют под названием задний мозг. Белое вещество моста в его передней части представлено поперечно идущими волокнами, направляющимися в средние ножки мозжечка. Они пронизываются продольными пучками волокон пирамидных путей, образующих затем пирамиды продолговатого мозга и направляющиеся в спинной мозг.
В продолговатом мозге и мосту основная часть серого вещества располагается в белом отдельными группами, или ядрами, которые являются рефлекторными центрами. Из 12 пар черепных (черепно-мозговых) нервов, выходящих из головного мозга и черепа самостоятельно, без связи со спинным мозгом, V-XII пары отходят от ядер, расположенных в продолговатом мозге и мосту (рис. 32). Часть из этих нервов иннервируют лицевую и глазодвигательную мускулатуру, что свидетельствует о сегментарных чертах строения продолговатого мозга и моста.
Функции продолговатого мозга и варолиева моста
Продолговатый мозг и мост выполняют проводниковую, рефлекторную и интегративную функции. В них поступают афферентные нервные волокна от вестибулярных и слуховых рецепторов, от кожи и мышц головы, от внутренних органов. Проводниковая функция продолговатого мозга и варолиева моста заключается в передаче импульсов из спинного мозга в вышележащие отделы ЦНС и в проведении импульсов, идущих из головного в спинной мозг. В продолговатом мозге заканчивается кортико-бульбарный нисходящий путь, по которому импульсы от коры больших полушарий поступают к ядрам черепно-мозговых нервов. От продолговатого мозга к спинному отходят нисходящие ретикуло-спинальные пути и вестибуло-спинальный пучок.
Через продолговатый мозг проходят восходящие пути от рецепторов слуховой и вестибулярной чувствительности. В продолговатом мозге афферентные нервные волокна, несущие информацию от рецепторов кожи и мышечных рецепторов, образуют синапсы на вторых нейронах, аксоны которых составляют путь в таламус.
Продолговатый мозг и мост выполняют сложные рефлекторные функции, в том числе и жизненно важные для организма. На дне IV-го желудочка (рис. 33) в ретикулярной формации продолговатого мозга на-
ходятся жизненно важные дыхательный центр, регулирующий дыхание, и сосудодвигательный центр, регулирующий работу сердца и просвет кровеносных сосудов. В продолговатом мозге и мосту находится ряд рефлекторных центров, связанных с процессами пищеварения, а именно: центры жевания, сосания, слюноотделения, глотания. Здесь же находятся центры некоторых защитных рефлексов: чихания, кашля, рвоты, мигания, слёзоотделения.
 Целая группа рефлексов продолговатого мозга и моста направлена на поддержание позы тела. В зависимости от рецепторов, при возбуждении которых осуществляется поддержание позы, эти рефлексы подразделяют на шейные тонические рефлексы и вестибулярные (лабиринтные). Шейные тонические рефлексы запускаются при возбуждении проприорецепторов мышц шеи вследствие изменения положения головы относительно туловища.
Целая группа рефлексов продолговатого мозга и моста направлена на поддержание позы тела. В зависимости от рецепторов, при возбуждении которых осуществляется поддержание позы, эти рефлексы подразделяют на шейные тонические рефлексы и вестибулярные (лабиринтные). Шейные тонические рефлексы запускаются при возбуждении проприорецепторов мышц шеи вследствие изменения положения головы относительно туловища.
Вестибулярные рефлексы неразрывно связаны с шейными тоническими рефлексами и дополняют их, т.к. не зависят от положения головы относительно туловища. Вестибулярные рефлексы подразделяются на статические и статокинетические. Статические рефлексы вызываются главным образом возбуждением рецепторов, расположенных в мешочке и маточке вестибулярного аппарата, и обеспечивают поддержание позы и равновесия тела при разнообразных неподвижных его положениях в пространстве. Среди статических рефлексов различают две группы: 1) позно-тонические рефлексы, или рефлексы положения, благодаря которым сохраняется определённая поза; 2) установочные, или выпрямительные рефлексы, которые обеспечивают переход из неестественной позы в обычное положение.
Статокинетические рефлексы обеспечивают поддержание позы тела при изменении скорости движения. Эти рефлексы вызываются при возбуждении рецепторов, расположенных в полукружных каналах вестибулярного аппарата. Возбуждение же рецепторов возникает при движении эндолимфы в каком-либо из трёх полукружных каналов в результате ус-
корения или торможения тела человека. К статокинетическим вестибулярным рефлексам относятся также лифтные рефлексы, которые проявляются в увеличении тонуса мышц сгибателей и уменьшении тонуса мышц разгибателей при ускорении вверх. При ускорении вниз наблюдается обратная картина: тонус мышц сгибателей уменьшается, а тонус мышц разгибателей повышается. В результате в начале подъёма происходит сгибание конечностей и человек слегка приседает, а в начале спуска – их разгибание и человек слегка выпрямляется.
СРЕДНИЙ МОЗГ
Строение среднего мозга
Средний мозг совместно с варолиевым мостом и продолговатым мозгом образует ствол головного мозга, в котором он занимает относительно небольшую по объёму часть. К стволу мозга иногда также относят промежуточный мозг и мозжечок. Средний мозг расположен над варолиевым мостом спереди от мозжечка. Он состоит из ножек мозга и крыши среднего мозга, или четверохолмия (рис. 34).
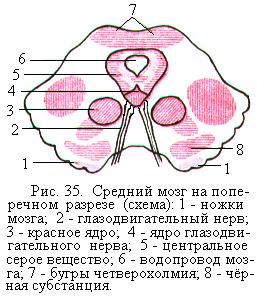
 Ножки мозга занимают переднюю часть среднего мозга и представляют собой белые округлые тяжи, выходящие из моста и направляющиеся к большим полушариям. На поперечном разрезе среднего мозга видно, что каждая ножка состоит из покрышки и основания, на границе которых находится чёрное вещество, или чёрная субстанция. Чёрная субстанция представляет собой скопление тел нейронов, содержащих пигмент меланин, придающий нейронам характерный тёмный цвет.
Ножки мозга занимают переднюю часть среднего мозга и представляют собой белые округлые тяжи, выходящие из моста и направляющиеся к большим полушариям. На поперечном разрезе среднего мозга видно, что каждая ножка состоит из покрышки и основания, на границе которых находится чёрное вещество, или чёрная субстанция. Чёрная субстанция представляет собой скопление тел нейронов, содержащих пигмент меланин, придающий нейронам характерный тёмный цвет.
Основание ножек образовано в основном нервными волокнами, идущими из коры больших полушарий в спинной, продолговатый мозг и мост. Покрышка ножек мозга состоит из восходящих нервных волокон, направляющихся в отдел промежуточного мозга – таламус. Среди этих волокон залегают ядра, самыми крупными из которых являются красные ядра. Они представляют собой удлинённые образования, расположенные между чёрной субстанцией и находящимся в центре серым веществом (рис. 35).
Четверохолмие составляет заднюю часть среднего мозга. Перпендикулярными бороздами четверохолмие делится на верхние, или передние, и нижние, или задние холмики (двухолмия), или бугры. В углублении между верхними холмиками находится железа внутренней секреции –
эпифиз. Между ножками мозга и четверохолмием в центре среднего мозга проходит узкий канал – водопровод мозга (Сильвиев водопровод), сообщающийся снизу с IV-м желудочком, а сверху – с III-м желудочком.
Функции среднего мозга
Средний мозг выполняет рефлекторную, интегративную и проводниковую функции. Он участвует в осуществлении целого ряда важных безусловных рефлексов. Так, при участии бугров четверохолмия осуществляются ориентировочные зрительные и слуховые рефлексы. Передние бугры четверохолмия являются первичными подкорковыми зрительными центрами, а задние бугры – первичными подкорковыми слуховыми центрами. Ядра, расположенные в передних и задних буграх четверохолмия, участвуют в осуществлении сторожевого рефлекса при внезапной подаче светового или звукового раздражителей. Этот рефлекс проявляется в перераспределении мышечного тонуса: происходит повышение тонуса мышц-сгибателей и уменьшение тонуса мышц-разгибателей, что мобилизует организм на быструю ответную реакцию.
 Чёрная субстанция среднего мозга участвует в регуляции тонуса мышц и различных тонких движений (например, мелких движений пальцев рук). Чёрная субстанция функционально связана с подкорковыми ядрами больших полушарий – полосатым телом и бледным шаром. Нейроны чёрной субстанции имеют дофаминергическую природу, т.к. синтезируют медиатор дофамин. Аксоны этих нейронов идут к полосатому телу и образуют синапсы на его нервных клетках, также содержащих в значительных количествах дофамин. Повреждение чёрной субстанции приводит к дегенерации дофаминергических нервных путей к полосатому телу, что является одной из причин возникновения тяжёлого заболевания нервной системы – болезни Паркинсона.
Чёрная субстанция среднего мозга участвует в регуляции тонуса мышц и различных тонких движений (например, мелких движений пальцев рук). Чёрная субстанция функционально связана с подкорковыми ядрами больших полушарий – полосатым телом и бледным шаром. Нейроны чёрной субстанции имеют дофаминергическую природу, т.к. синтезируют медиатор дофамин. Аксоны этих нейронов идут к полосатому телу и образуют синапсы на его нервных клетках, также содержащих в значительных количествах дофамин. Повреждение чёрной субстанции приводит к дегенерации дофаминергических нервных путей к полосатому телу, что является одной из причин возникновения тяжёлого заболевания нервной системы – болезни Паркинсона.
В покрышке среднего мозга располагаются ядра глазодвигательного (рис. 35) и блокового нервов (соответственно III-й и IV-й пары черепномозговых нервов). Эти нервы управляют движениями глаз, аккомодацией глаза, зрачковым рефлексом.
Важным промежуточным центром проводящих путей ствола мозга являются красные ядра. Они связаны нервными путями с корой больших полушарий, ретикулярной формацией, мозжечком и спинным мозгом. От красных ядер берёт начало нисходящий руброспинальный путь, идущий к мотонейронам спинного мозга. С его помощью осуществляется регуляция тонуса скелетных мышц, а именно: происходит повышение тонуса мышц-
сгибателей и уменьшение тонуса мышц-разгибателей. Это имеет большое значение при поддержании позы и осуществлении движений. Если у жи  вотного перерезать головной мозг на уровне задних бугров четверохолмия так, чтобы красные ядра оказались выше линии перерезки, то у такого животного наступает резкое повышение тонуса разгибательной мускулатуры (рис. 36). Конечности у него сильно выпрямляются, голова запрокинута назад, хвост приподнят, конечности трудно согнуть даже силой. Животное не может принять нормальную позу. Такое состояние мускулатуры тела называется децеребрационной ригидностью.
вотного перерезать головной мозг на уровне задних бугров четверохолмия так, чтобы красные ядра оказались выше линии перерезки, то у такого животного наступает резкое повышение тонуса разгибательной мускулатуры (рис. 36). Конечности у него сильно выпрямляются, голова запрокинута назад, хвост приподнят, конечности трудно согнуть даже силой. Животное не может принять нормальную позу. Такое состояние мускулатуры тела называется децеребрационной ригидностью.
Предполагается, что красные ядра являются не только источником, но и посредником в регуляции разгибательных рефлексов, передавая тормозные влияния от мозжечка и моторных областей коры. Удаление мозжечка или моторных областей коры приводит к усилению тонуса мышц-разгибателей.
РЕТИКУЛЯРНАЯ ФОРМАЦИЯ
Ретикулярная формация начинается в срединной части верхних шейных сегментов спинного мозга и продолжается в центральных отделах продолговатого мозга, моста, среднего и промежуточного мозга. Она представляет собой скопления нейронов (ядра) с многочисленными сильно ветвящимися отростками, идущими в разных направлениях и образующими густую сеть. Между нейронами ретикулярной формации образуется очень много синапсов.
В ретикулярной формации человека выделяют 48 ядер. Наиболее крупным является гигантоклеточное ядро, которое содержит гигантские нейроны, отсутствующие в других ядрах. Важным свойством нейронов ретикулярной формации является их высокая химическая чувствительность к различным гуморальным факторам и фармакологическим веществам, особенно к анестезирующим препаратам и так называемым успокаивающим средствам.
В ретикулярной формации продолговатого мозга располагаются жизненно важные центры регуляции дыхания, сердечно-сосудистой системы, деятельности пищеварительного тракта, центры рефлекторных актов, связанных с вестибулярными и слуховыми нервами.
Установлено, что ретикулярная формация по восходящим нервным путям оказывает возбуждающее влияние на кору больших полушарий, а по нисходящим путям - возбуждающее или тормозящее действие на деятельность спинного мозга. Было обнаружено, что электрическое раздражение гигантоклеточного ядра ретикулярной формации вызывает неспецифическое торможение сгибательных и разгибательных рефлексов, осуществляемых мотонейронами спинного мозга. Влияние ретикулярной формации на мышечный тонус передаётся по двум ретикуло-спинальным путям: быстропроводящему и медленнопроводящему. Импульсы, поступающие по этим путям, повышают активность гамма-мотонейронов спинного мозга, что в свою очередь возбуждает альфа-мотонейроны, и мышечный тонус увеличивается. Ретикулярная формация может выступать не только в роли регулятора возбудимости мотонейронов спинного мозга, но и принимать участие в процессах, связанных с поддержанием позы и организацией целенаправленных движений.
|
|
|
|
|
Дата добавления: 2014-12-26; Просмотров: 1314; Нарушение авторских прав?; Мы поможем в написании вашей работы!