КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Строение синапса. Синаптическая передача возбуждения ….….. 23 7 страница
|
|
|
|
Если человек из состояния покоя переходит ко сну, то в его ЭЭГ появляются более медленные и высокоамплитудные волны, образующие тета-ритм (θ) с частотой 4-7 колебаний в 1 секунду и амплитудой 100-150 мкВ. Тета-ритм регистрируется также при отрицательных эмоциях, болевых раздражениях, небольшом наркозе.
Во время глубокого сна, глубокого наркоза, при потере сознания и некоторых патологических состояниях коры наблюдается дельта-ритм (δ), имеющий частоту 0,5-3,5 колебаний в 1 секунду и амплитуду 250-300 мкВ (рис. 54).
На характер ЭЭГ влияют самые различные факторы. Например, при ухудшении кровоснабжения мозга электрическая активность мозга понижается, а при уменьшении содержания СО2 в крови в ЭЭГ наблюдается увеличение низкочастотных колебаний.
Для исследования функций коры больших полушарий кроме ЭЭГ применяется также метод вызванных потенциалов. Вызванные потенциалы представляют собой регистрируемые с поверхности коры или кожи
головы электрические колебания в ответ на раздражение рецепторов, периферических нервов, ядер таламуса и других структур, участвующих в проведении сенсорных сигналов.
 Вызванные потенциалы разделяют на две группы: первичные ответы и вторичные ответы. Первичный ответ представляет собой двухфазное колебание потенциала, складывающееся из позитивной и негативной волны. Первичные ответы регистрируются только в проекционных зонах и обозначаются числом, которое равно отрезку времени в миллисекундах от начала раздражения рецепторов или нервов до появления вызванного потенциала.
Вызванные потенциалы разделяют на две группы: первичные ответы и вторичные ответы. Первичный ответ представляет собой двухфазное колебание потенциала, складывающееся из позитивной и негативной волны. Первичные ответы регистрируются только в проекционных зонах и обозначаются числом, которое равно отрезку времени в миллисекундах от начала раздражения рецепторов или нервов до появления вызванного потенциала.
Предполагается, что начальная позитивная фаза первичных ответов обусловлена суммацией синаптических потенциалов тел пирамидных нейронов и тех дендритов, которые отходят от тел пирамидных нейронов вглубь коры. Следующая за позитивной негативная фаза первичного ответа вызвана активацией тех дендритов пирамидных нейронов, которые расположены в поверхностных слоях коры.
Вторичные ответы, как правило, регистрируются в ассоциативных зонах коры, примыкающих к проекционным. Они отражают процессы, связанные с передачей возбуждения от первичных проекционных зон к ассоциативным и с рабочей настройкой нейронов коры путём изменения их возбудимости.
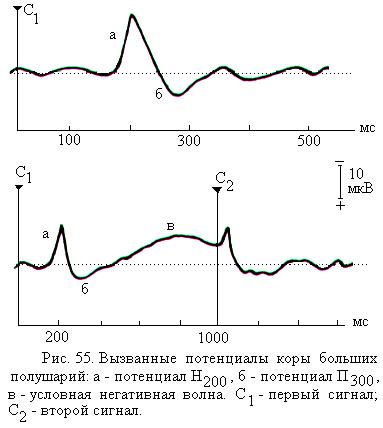
Выявлена связь некоторых отдельных колебаний вызванных потенциалов с такими процессами как внимание, обработка поступившей информации, ожидание подачи сигнала. Например, обработка слуховой информации ведёт к появлению позитивной волны П300, концентрация внимания отражается на параметрах негативной во-лны Н200 (рис. 55).
Специфические вызванные потенциалы мо-жно зарегистрировать при формировании условного рефлекса. Примером таких потенциалов может служить описанная Г. Уолтером волна ожидания, или условная негативная волна, или Е-волна. Так, если первый
условный сигнал (например, команда «внимание») информирует о том, что человек должен будет совершить движение при действии последующего второго раздражителя (например, команды «марш»), то после ряда сочетаний первый раздражитель начинает вызывать волну ожидания, которая продолжается до подачи второго раздражителя (рис. 55, в). Волна ожидания представляет собой медленное негативное колебание с амплитудой до 40 мкВ. Волна ожидания предшествует движению, которое должен произвести субъект, и, по-видимому, является электрофизиологическим отражением процессов подготовки к целенаправленному движению. Однако более полно связь волны ожидания с психическими процессами у человека пока не расшифрована.
Функциональная асимметрия больших полушарий
 Конечный мозг у человека образован правым и левым полушариями, каждое из которых состоит из морфологически почти одинаковых долей. Подобно тому, как существует специализация различных долей каждого полушария, так и два полушария играют разную роль в осуществлении высших нервных функций.
Конечный мозг у человека образован правым и левым полушариями, каждое из которых состоит из морфологически почти одинаковых долей. Подобно тому, как существует специализация различных долей каждого полушария, так и два полушария играют разную роль в осуществлении высших нервных функций.
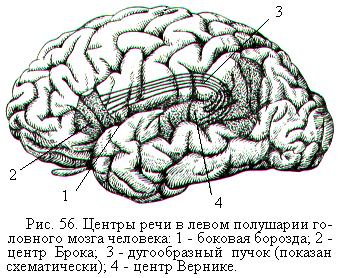
После открытия во второй половине 19-го века французским учёным Полем Брокá моторного центра речи (центр Брока) и немецким учёным-невропатологом Карлом Вернике сенсорного центра речи (центр Вернике), расположенных в левом полушарии (рис. 56), это полушарие стали считать главным в отношении речевой функции и мышления. Центр Брока находится в задней части нижней лобной извилины, он обеспечивает способность человека говорить. Центр Вернике расположен в задней части верхней височной извилины. Благодаря нему человек понимает свою и чужую речь. Центр Вернике и центр Брока связаны проходящим под корой пучком нервных волокон, который получил название дугообразного пучка.
После обнаружения центров речи долгое время не появлялось никаких новых данных о неравнозначности полушарий. Речь казалась единственным исключением из общего правила, согласно которому оба полушария совершенно равноценны в отношении всех остальных сенсорных и двигательных функций. Только в 50-х годах 20-го века Роджер Сперри с сотрудниками провели ряд экспериментов с перерезкой у кошек мозолистого тела – толстого пучка, содержащего миллионы нервных волокон и
соединяющего оба полушария. Когда таким животным помещали разные объекты перед правой и левой половиной поля зрения, то оказалось, что правое и левое полушарие работают независимо друг от друга, т.е. зрительная информация не передавалась от одного полушария к другому.
У людей, больных эпилепсией, во время припадка бурная импульсная активность нейронов распространяется от поражённого участка на другие области мозга. Она может через мозолистое тело передаться другому полушарию и человек может погибнуть. Чтобы сдержать нервный взрыв и спасти больного, нейрохирурги перерезают мозолистое тело. После такой операции у больных не отмечается практически никаких видимых изменений в отношении свойств личности, интеллекта или поведения.
 Однако, как показали исследования Р. Сперри, за которые в 1981 г. ему была присуждена Нобелевская премия, если предъявлять зрительную информацию каждому полушарию в отдельности, то полушария будут функционировать и научаться независимо друг от друга.
Однако, как показали исследования Р. Сперри, за которые в 1981 г. ему была присуждена Нобелевская премия, если предъявлять зрительную информацию каждому полушарию в отдельности, то полушария будут функционировать и научаться независимо друг от друга.
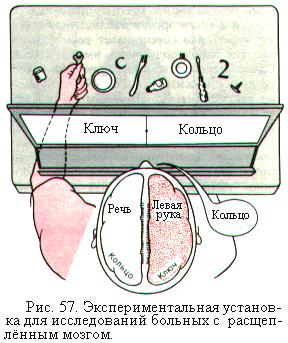
Так, если в левую половину поля зрения предъявить испытуемому слово ключ, то информация о нём поступит в правое полушарие. Испытуемый не может назвать это слово, но левой рукой выбирает среди предметов ключ (рис. 57). Если же в правую половину поля зрения предъявить слово кольцо, информация о котором поступит в левое полушарие, то испытуемый назовёт это слово, но выбрать его на ощупь, без контроля зрения, среди других предметов не может.
После исследований Р. Сперри деятельности мозга у людей с перерезанными связями между полушариями («расщеплённый мозг») стало складываться представление о частичном доминировании полушарий мозга человека, или о функциональной межполушарной асимметрии.
Было обнаружено существование 5-и основных различий полушарного доминирования у человека: речевое – неречевое, время – пространство, анализ – синтез, последовательное восприятие – одновременное восприятие, абстрактное восприятие – конкретное восприятие.
Выделяют моторную, сенсорную и психическую асимметрию. Моторной асимметрией называют неодинаковость двигательной активности рук, ног, лица, половин тела, управляемой каждым полушарием мозга. Сенсорной асимметрией обозначают неравнозначность восприятия каждым из полушарий объектов, расположенных слева и справа от средней плоскости тела. Психической асимметрией называют специализацию полушарий мозга в отношении различных форм психической деятельности.
Предполагают, что в процессе обучения правое полушарие работает по принципу дедукции, т.е. сначала осуществляет синтез, а затем анализ. Левое же полушарие функционирует по принципу индукции, сначала анализируя раздражители, а затем синтезируя их.
Было установлено, что левое полушарие доминирует в отношении речи, сложных произвольных движений, чтения, письма и счёта, в оценке времени, установлении сходства. Правое же полушарие ответственно в основном за неречевые функции – распознавание сложных зрительных и слуховых образов, тактильное восприятие, восприятие пространства, формы и направления, установление различий, за интуицию.
На основании результатов изучения расщеплённого мозга было обнаружено, что левое полушарие осуществляет переработку информации аналитически и последовательно, а правое – одновременно и целостно.
Более чем у 95% всех праворуких людей, не имевших в раннем возрасте травм или поражений мозга, язык и речь контролируется левым полушарием, а у остальных 5% - правым. Большая часть леворуких людей, около 70%, также имеют речевые зоны в левом полушарии. У половины из остальных левшей (примерно 15%) речь контролируется одним правым полушарием, а у другой половины (примерно 15%) – обоими полушариями.
Предполагается, что у мужчин функциональная асимметрия полушарий выражена в большей степени, чем у женщин. У мужчин поражение левого полушария вследствие инсульта или опухоли мозга вызывало потерю речи в три раза чаще, чем у женщин, или приводило к гораздо большему ухудшению речевых способностей. У мужчин поражение левого полушария ухудшало результаты речевого теста в большей степени, чем неречевого (невербального). При поражении правого полушария получалось обратное соотношение. Для женщин сторона, на которой произошло поражение, не имела столь большого значения. Следовательно, у мужчин специализация полушарий, по-видимому, выражена больше, чем у женщин.
ПЕРИФЕРИЧЕСКАЯ НЕРВНАЯ СИСТЕМА
Отделы нервной системы. Соматическая нервная система
Нервную систему делят на отделы по анатомическому и функциональному признакам. Анатомически она подразделяется на центральную нервную систему и периферическую нервную систему, функционально – на вегетативную (автономную) и соматическую. Вегетативная нервная система регулирует обмен веществ, работу внутренних органов и гладких мышц. Соматической нервной системой называют отдел нервной системы, иннервирующий скелетную мускулатуру и кожу и обеспечивающий движение организма, а также его чувствительность к условиям внешней среды.
Соматические компоненты реакций организма, осуществляемые скелетной мускулатурой, в отличие от вегетативных могут быть по желанию человека (т.е. произвольно) вызваны, усилены или заторможены. Они находятся в течение всего хода реакции под контролем сознания. Соматическая нервная система имеет характерные особенности, отличающие её от вегетативной нервной системы.
Первая особенность заключается в расположении эфферентного нейрона. В соматической нервной системе вставочный нейрон и тело эфферентного нейрона, принимающие участие в осуществлении рефлекса, находятся в пределах ЦНС, т.е. в сером веществе спинного или головного мозга.
Вторая особенность относится к иннервации органов, снабжаемых волокнами соматической нервной системы. Перерезка передних корешков спинного мозга сопровождается полным отмиранием всех эфферентных соматических волокон, т.к. они отделяются от тел нейронов, залегающих в передних рогах спинного мозга.
Третья особенность касается выхода и распределения нервных волокон. Соматические нервные волокна покидают ствол головного мозга и спинной мозг сегментарно и перекрывают иннервируемые области не менее трёх смежных сегментов. На периферии соматические волокна распределены строго сегментарно.
Четвёртая особенность состоит в морфологических и функциональных параметрах волокон. Соматические эфферентные нервные волокна толстые, миелинизированные, легко возбудимы и обладают высокой скоростью проведения возбуждения.
Закономерности строения периферической нервной системы
Периферическая нервная система образована нервными узлами (ганглиями), нервными сплетениями, нервами и нервными окончаниями, расположенными за пределами ЦНС. В зависимости от выполняемой функции различают нервы чувствительные, двигательные и смешанные. Чувствительные нервы сформированы отростками нейронов чувствительных узлов черепномозговых нервов или спинномозговых узлов. Двигательные нервы состоят из аксонов нейронов, тела которых расположены в двигательных ядрах черепномозговых нервов или в передних рогах спинного мозга.
Кроме соматических нервов, в состав периферической нервной системы входят вегетативные нервы, образованные отростками клеток вегетативных ядер черепномозговых нервов или нейронов, тела которых лежат в боковых рогах спинного мозга. В периферической нервной системе
человека преобладают смешанные нервы, содержащие и чувствительные и двигательные волокна.
В строении периферической нервной системы имеется ряд закономерностей. Основными из них являются следующие: 1) нервы являются парными и расходятся симметрично в стороны от головного и спинного мозга, лежащего по осевой линии тела; 2) нервы идут к органам по кратчайшему пути, удлиняясь и следуя за органами, если они перемещаются в процессе внутриутробного развития; 3) нервы, иннервирующие мышцы, отходят от тех сегментов спинного мозга, которые соответствуют миотомам, из которых происходят эти мышцы; 4) нервные стволы сопровождают артерии, вены, лимфатические сосуды, образуя сосудисто-нервные пучки.
Черепномозговые нервы
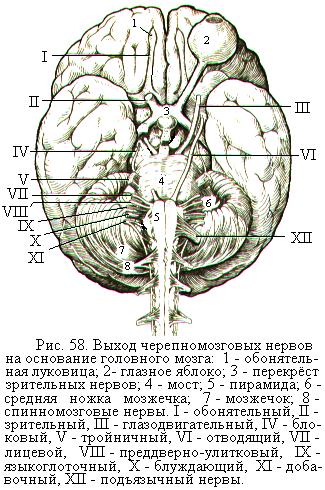 Нервные стволы (нервы) представляют собой объединение многочисленных нервных волокон сразу после выхода их из мозга. Эту начальную часть нерва называют корешком. От головного мозга отходит 12 пар черепномозговых, или черепных, нервов (рис. 58). Каждая пара нервов имеет свой номер и название. Часть черепномозговых нервов являются только двигательными (III, IV, VI, XI и XII пары), часть – чувствительными (I, II, VIII пары), а остальные (V, VII, IX и X пары) – смешанные. В состав некоторых черепномозговых нервов (III, VII, IX и X пары) входят также парасимпатические волокна, иннервирующие железы и гладкие мышцы.
Нервные стволы (нервы) представляют собой объединение многочисленных нервных волокон сразу после выхода их из мозга. Эту начальную часть нерва называют корешком. От головного мозга отходит 12 пар черепномозговых, или черепных, нервов (рис. 58). Каждая пара нервов имеет свой номер и название. Часть черепномозговых нервов являются только двигательными (III, IV, VI, XI и XII пары), часть – чувствительными (I, II, VIII пары), а остальные (V, VII, IX и X пары) – смешанные. В состав некоторых черепномозговых нервов (III, VII, IX и X пары) входят также парасимпатические волокна, иннервирующие железы и гладкие мышцы.
I пара – обонятельный нерв, образован отростками обонятельных рецепторных клеток, расположенных в слизистой оболочке носовой полости, идёт к обонятельным луковицам переднего мозга.
II пара – зрительный нерв, образован отростками нервных клеток сетчатки глаза. Войдя из глазницы в полость черепа, правый и левый зрительные нервы частично перекрещиваются (зрительный перекрёст) и продолжаются в зрительные тракты.
III пара – глазодвигательный нерв, иннервирует верхнюю, нижнюю и медиальную прямые мышцы глазного яблока, нижнюю косую мышцу глаза и мышцу, поднимающую верхнее веко. Входящие в состав глазодвигательного нерва парасимпатические волокна иннервируют мышцу, суживающую зрачок, и ресничную мышцу, способствующую изменению кривизны хрусталика.
IV пара – блоковой нерв, иннервирует верхнюю косую мышцу глаза.
V пара – тройничный нерв, выходит из мозга двумя корешками: двигательным (меньшим) и чувствительным (бóльшим). Волокна тройничного нерва образуют три ветви: глазной нерв, верхнечелюстной нерв и нижнечелюстной нерв. Чувствительные волокна тройничного нерва иннервируют кожу лица, переднюю часть головы, глаза, слизистые оболочки носовой и ротовой полостей, придаточных пазух носа. Двигательные волокна тройничного нерва иннервируют жевательные мышцы, мышцы дна рта, мышцу натягивающую мягкое нёбо и одну из мышц барабанной полости среднего уха.
VI пара – отводящий нерв, иннервирует наружную прямую мышцу глаза, которая отводит (поворачивает) глазное яблоко вбок.
VII пара – лицевой нерв. Двигательные его волокна иннервируют мимические мышцы лица и некоторые мышцы шеи. Чувствительные волокна, входящие в состав лицевого нерва под названием промежуточный нерв, иннервируют слизистую оболочку передних 2/3 языка, железы слизистой оболочки носовой и ротовой полостей, подъязычные и подчелюстные слюнные железы, слёзные железы.
VIII пара – преддверно-улитковый нерв, идёт от внутреннего уха. Состоит из отростков нейронов улиткового узла, несущих информацию от рецепторов слухового аппарата, и из отростков нейронов преддверного узла, проводящих импульсы от рецепторов вестибулярного аппарата.
IX пара – языкоглоточный нерв, содержит двигательные, чувствительные и парасимпатические волокна. Двигательные его волокна участвуют в иннервации мышц глотки. Чувствительные волокна иннервируют слизистую оболочку задней трети языка, глотки, среднего уха. Парасимпатические волокна иннервируют слюнные околоушные железы.
X пара – блуждающий нерв, самый длинный из черепномозговых нервов. Снабжает своими ветвями органы дыхания, бóльшую часть пищеварительного тракта, доходя до ободочной кишки, печень, поджелудочную железу, почки, а также даёт ветви к сердцу и начальной части аорты. Двигательные волокна блуждающих нервов иннервируют поперечнополосатые мышцы глотки, мягкого нёба и гортани; парасимпатические – гладкую мускулатуру внутренних органов, сердечную мышцу и секреторные железы пищеварительного тракта; чувствительные волокна проводят импульсы от рецепторов внутренних органов, заднего отдела твёрдой оболочки головного мозга и от наружного уха.
XI пара – добавочный нерв, иннервирует грудино-ключично-сосцевидную и трапециевидную мышцы.
XII пара – подъязычный нерв, иннервирует мышцы языка. В составе подъязычного нерва проходит часть двигательных волокон от 1-го спинномозгового нерва, которые иннервируют некоторые мышцы шеи.
Спинномозговые нервы
У человека имеется 31 пара спинномозговых нервов. Они отходят от спинного мозга двумя корешками: передним, состоящим из двигательных волокон, и задним, состоящим из чувствительных волокон. В области межпозвоночного отверстия они соединяются в один ствол – смешанный спинномозговой нерв. Перед местом соединения задний корешок образует нервный спинномозговой узел, состоящий из тел чувствительных нейронов с Т-образно ветвящимися отростками. Длинный отросток в данном случае является дендритом и направляется на периферию, где заканчивается рецептором, а короткий отросток, служащий аксоном, в составе заднего корешка входит в задние рога спинного мозга.
Спинномозговые нервы бывают различной толщины и длины. Чем крупнее нервный ствол, тем большую область тела он иннервирует. Поэтому по толщине, длине, положению и ветвлению спинномозгового нерва можно приблизительно судить об области, иннервируемой им. Самые длинные – это нервные стволы конечностей, особенно нижних.
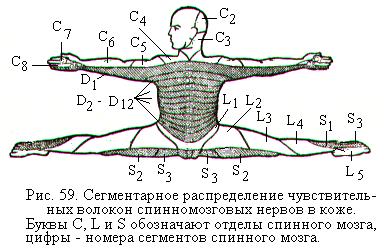 Спинномозговые нервы отходят от спинного мозга в обе стороны в правильном порядке так, что можно выделить подобные друг другу участки - сегменты. Спинномозговые нервы соответствуют сегментам, или метамерам, тела и сегментам спинного мозга, поэтому обозначаются латинскими буквами соответственно сегментам спинного мозга, из которого выходят корешки нерва (например, С1, L4). Каждый сегмент спинного мозга связан спинномозговым нервом с сегментом тела (рис. 59). Эта связь сохраняется, начиная с эмбрионального периода, на протяжении всей жизни человека.
Спинномозговые нервы отходят от спинного мозга в обе стороны в правильном порядке так, что можно выделить подобные друг другу участки - сегменты. Спинномозговые нервы соответствуют сегментам, или метамерам, тела и сегментам спинного мозга, поэтому обозначаются латинскими буквами соответственно сегментам спинного мозга, из которого выходят корешки нерва (например, С1, L4). Каждый сегмент спинного мозга связан спинномозговым нервом с сегментом тела (рис. 59). Эта связь сохраняется, начиная с эмбрионального периода, на протяжении всей жизни человека.
В процессе развития вместе с каждым нервом связан участок развивающихся мышц (миотом) и кожи (дерматом). Задний корешок и спинномозговой узел связаны с сегментом кожи (рис. 59), они сформировались из соответствующего дерматома. Передний корешок связан с мышцами, которые произошли из соответствующего миотома. На основании этого выделяют сегментарную иннервацию мышц и кожи. Вместе с тем для надёжности каждый сегмент тела иннервируется не только одним спинномозговым нервом из своего сегмента спинного мозга, а также из выше- и нижележащего сегмента. Периферические разветвления спинномозговых нервов не соблюдают такой правильности распределения. У человека из 31 пары спинномозговых нервов 8 шейных (С), 12 грудных (Th), 5 поясничных (L), 5 крестцовых (S) и 1 копчиковая пара (Co).
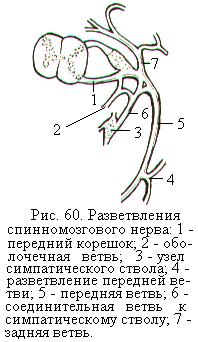 Каждый спинномозговой нерв после выхода из межпозвоночного отверстия делится на четыре ветви: 1) переднюю, иннервирующую переднюю стенку туловища и конечности; 2) заднюю для иннервации мышц и кожи спины и затылка; 3) соединительную, идущую к узлу симпатического ствола; 4) менингеальную (оболочечную), направляющуюся обратно в позвоночный канал для иннервации оболочек спинного мозга (рис. 60).
Каждый спинномозговой нерв после выхода из межпозвоночного отверстия делится на четыре ветви: 1) переднюю, иннервирующую переднюю стенку туловища и конечности; 2) заднюю для иннервации мышц и кожи спины и затылка; 3) соединительную, идущую к узлу симпатического ствола; 4) менингеальную (оболочечную), направляющуюся обратно в позвоночный канал для иннервации оболочек спинного мозга (рис. 60).
Задние ветви сохраняют сегментарный принцип иннервации и иннервируют кожу затылочной области, кожу и мышцы задней области шеи, спины, поясничной области и ягодиц. Передние ветви спинномозговых нервов иннервируют кожу и мышцы передней и боковых частей шеи, груди, живота, конечностей (рис. 59). Они сохраняют сегментарность иннервации только в грудном отделе (межреберные нервы). В остальных отделах передние ветви спинномозговых нервов соединяются друг с другом петлями, образуя шейное, плечевое, поясничное, крестцовое и копчиковое сплетения, от которых отходят периферические нервы.
Шейное сплетение образовано передними ветвями четырёх верхних шейных спинномозговых нервов. От него отходят: 1) чувствительные (кожные) нервы, иннервирующие кожу затылочной области, ушной раковины, наружного слухового прохода, шеи, верхней части груди и спины; 2) двигательные нервы к близлежащим мышцам шеи; 3) смешанный диафрагмальный нерв.
Плечевое сплетение образовано передними ветвями V – VIII-го шейных и частично I-го грудного спинномозговых нервов. От сплетения отходит много длинных и коротких нервов, иннервирующих кожу, мышцы и суставы верхних конечностей, часть мышц шеи, груди и спины.
Передние ветви 12 пар грудных спинномозговых нервов сплетения не образуют и идут в межреберных промежутках. Они называются межреберными нервами и иннервируют мышцы груди и мышцы передней брюшной стенки. Передние и боковые кожные ветви этих нервов иннервируют кожу груди и живота.
Поясничное сплетение образовано передними ветвями I – III поясничных и частично XII-го грудного и IV-го поясничного спинномозговых нервов. Чувствительные нервы, выходящие из этого сплетения, иннервируют кожу нижней части передней брюшной стенки и частично бедра, голени и стопы, наружных половых органов. Мышечные ветви иннервируют мышцы стенок живота и часть мышц бедра.
Крестцовое сплетение - самое крупное. Оно образовано передними ветвями I – IV крестцовых, V-го поясничного и частично IV-го поясничного спинномозговых нервов. Его нервы иннервируют мышцы и частично кожу ягодичной области и промежности, кожу наружных половых органов, кожу и мышцы задней поверхности бедра, кости, суставы, мышцы и кожу голени и стопы.
Копчиковое сплетение образовано передними ветвями V крестцового и I копчикового нервов. Его ветви иннервируют кожу в области копчика и в окружности заднего прохода.
ВЕГЕТАТИВНАЯ НЕРВНАЯ СИСТЕМА
Строение вегетативной нервной системы
Вегетативной нервной системой (ВНС) называют часть нервной системы, которая регулирует и координирует деятельность внутренних органов, обмен веществ, гладкую мускулатуру, железы внутренней секреции, постоянство внутренней среды организма и функциональную активность тканей. ВНС иннервирует весь организм, все органы и ткани. Структурные и функциональные особенности ВНС дали определённые основания рассматривать её как «автономную», т.е. не зависящую в своих функциях от деятельности центральной нервной системы и от воли человека. Однако представление об автономности вегетативной нервной системы является весьма условным. В настоящее время не подлежит сомнению, что посредством ВНС центральная нервная система регулирует функции внутренних органов, а также кровоснабжение и трофику всех тканей организма.
По анатомическому строению ВНС представляет совокупность следующих структурных образований: 1) нервных волокон; 2) периферических нервных узлов (ганглиев), состоящих из нервных клеток; 3) центров в сером веществе спинного, продолговатого и среднего мозга, от тел нейронов которых начинаются нервные волокна; 4) высших центров, находящихся в отделе промежуточного мозга – гипоталамусе.
Вегетативные рефлекторные дуги построены по такому же плану, что и соматические, и содержат чувствительные, вставочные и эфферентные звенья. Различие их заключается в том, что эфферентные нейроны ВНС лежат в ганглиях за пределами центральной нервной системы.
В вегетативной нервной системе выделяют симпатический отдел, или симпатическую нервную систему, и парасимпатический отдел, или
парасимпатическую нервную систему. Иногда выделяют ещё метасимпатическую часть ВНС. Сфера иннервации метасимпатической части ВНС охватывает только те внутренние органы, которые обладают собственным моторным ритмом, например желудок, кишечник.
Симпатический и парасимпатический отделы ВНС различаются между собой: 1) по расположению центров в мозге, от которых идут к органам нервные волокна; 2) по близости расположения ганглиев к органам-мишеням; 3) по медиатору, который используют постганглионарные нейроны в синапсах на клетках органов-мишеней для регулирования их функций; 4) по характеру оказываемых влияний на внутренние органы.
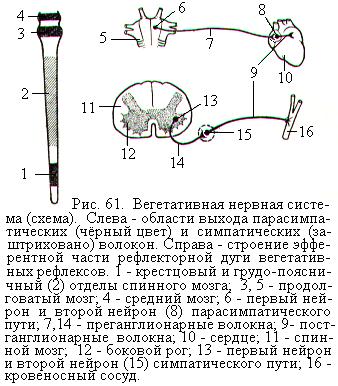 Центры ВНС, три из которых парасимпатические, а один симпатический, расположены в четырёх отделах головного и спинного мозга (рис. 61). К парасимпатическим центрам относятся: 1) мезэнцефальные центры, находящиеся в среднем мозге и посылающие вегетативные волокна в составе глазодвигательного нерва; 2) бульбарные центры, расположенные в продолговатом мозге и посылающие парасимпатические эфферентные волокна в составе лицевого, языкоглоточного и блуждающего нервов; 3) сакральные центры, залегающие в боковом промежуточном веществе II – IV-х крестцовых сегментов спинного мозга; волокна от них идут в составе тазовых нервов. К симпатическим центрам относятся тораколюмбальные центры, расположенные в боковых рогах I – XII-х грудных и I - II-х (иногда I – IV-х) поясничных сегментов спинного мозга. Вегетативные волокна от симпатических центров выходят через передние корешки спинномозговых нервов вместе с отростками мотонейронов.
Центры ВНС, три из которых парасимпатические, а один симпатический, расположены в четырёх отделах головного и спинного мозга (рис. 61). К парасимпатическим центрам относятся: 1) мезэнцефальные центры, находящиеся в среднем мозге и посылающие вегетативные волокна в составе глазодвигательного нерва; 2) бульбарные центры, расположенные в продолговатом мозге и посылающие парасимпатические эфферентные волокна в составе лицевого, языкоглоточного и блуждающего нервов; 3) сакральные центры, залегающие в боковом промежуточном веществе II – IV-х крестцовых сегментов спинного мозга; волокна от них идут в составе тазовых нервов. К симпатическим центрам относятся тораколюмбальные центры, расположенные в боковых рогах I – XII-х грудных и I - II-х (иногда I – IV-х) поясничных сегментов спинного мозга. Вегетативные волокна от симпатических центров выходят через передние корешки спинномозговых нервов вместе с отростками мотонейронов.
Периферическая часть ВНС образована выходящими из головного и спинного мозга вегетативными нервами, а также вегетативными сплетениями и их узлами, лежащими либо около позвоночника (паравертебральные узлы, или ганглии), либо в нервных сплетениях недалеко от внутренних органов (превертебральные ганглии), либо в стенках внутренних органов (интрамуральные ганглии). Одной из основных особенностей строения ВНС является то, что её эфферентные пути состоят из двух нейронов, в отличие от однонейронного эфферентного пути соматической нервной системы. Тела первых нейронов ВНС залегают в вегетативных центрах в головном или спинном мозге. Аксоны этих нейронов – преганглионарные волокна, покидая мозг, идут к вегетативным ганглиям, где и образуют синапсы на телах вторых эфферентных нейронов. Аксоны клеток вторых эфферентных нейронов называются постганглионарными волокнами, они идут к иннервируемому органу и образуют синапсы на его клетках (рис. 61).
Преганглионарные волокна ВНС относятся к типу В, имеют диаметр 2 – 3,5 мкм, покрыты тонкой миелиновой оболочкой и проводят импульсы со скоростью от 3 до 18 м в секунду. Постганглионарные волокна принадлежат к типу С, имеют диаметр до 2 мкм, большая часть их не покрыта миелиновой оболочкой. Скорость распространения по ним нервных импульсов от 1 до 3 м в секунду.
Симпатическая нервная система
Симпатическая нервная система состоит из центральной части, расположенной в боковых рогах от I-го грудного до II – IV-го поясничных сегментов спинного мозга, и периферической, включающей многочисленные нервы, узлы и сплетения. Симпатические нервные узлы подразделяются на две группы: паравертебральные (околопозвоночные) и превертебральные (предпозвоночные). Паравертебральные ганглии располагаются двумя цепочками по бокам от позвоночника и образуют правый и левый симпатические стволы. Превертебральные узлы находятся на достаточно большом отдалении как от позвоночника, так и от иннервируемых органов, и располагаются в периферических нервных сплетениях, лежащих в грудной и брюшной полостях. К числу наиболее крупных превертебральных ганглиев относятся чревный, верхний брыжеечный и нижний брыжеечный узлы. Два первых вместе с отходящими нервами образуют самое крупное из автономных сплетений организма – солнечное сплетение (рис. 62).
Симпатические нервные волокна выходят из спинного мозга в составе передних корешков спинномозговых нервов, а затем через соединительную ветвь направляются к соответствующему узлу симпатического ствола. Там бóльшая часть волокон образует синапсы на нейронах ганглия, аксоны которых (постганглионарные волокна) идут уже к органам. Другая, меньшая часть волокон следует через узел симпатического ствола без перерыва и подходит к превертебральным ганглиям. Там они (преганглионарные волокна) разветвляются и образуют синапсы на нейронах узла.
Преганглионарных волокон значительно меньше, чем ганглионарных нейронов. Так, к верхнему шейному симпатическому ганглию у человека подходит примерно 10 тысяч преганглионарных волокон, которые оканчиваются на миллионе нейронов ганглия. Это обеспечивает высокую надёжность проведения в ганглии и расширение зоны влияния преганглионарных волокон на иннервируемые органы. Аксоны ганглионарных нейронов (постганглионарные волокна) направляются к органам и синаптически контактируют с их клетками (рис. 62). Для постганглионарных симпатических волокон характерно образование сплетений по ходу артерий, снабжающих кровью данный орган.
|
|
|
|
|
Дата добавления: 2014-12-26; Просмотров: 548; Нарушение авторских прав?; Мы поможем в написании вашей работы!