КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Строение синапса. Синаптическая передача возбуждения ….….. 23 6 страница
|
|
|
|
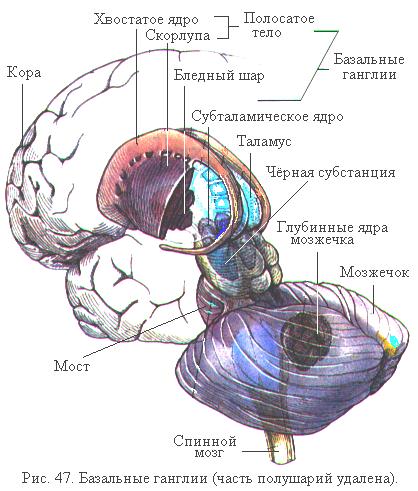
Хвостатое ядро, скорлупа и бледный шар формируют так называемую стриопаллидарную систему, к которой часто относят субталамическое ядро (люисово тело) промежуточного мозга и чёрную субстанцию среднего мозга (рис. 47). Все эти структуры функционируют совместно. Подкорковые ядра имеют многочисленные афферентные и эфферентные нервные связи с корой больших полушарий, средним и промежуточным мозгом, лимбической системой и мозжечком. Для нормального функционирования подкорковых ядер очень важное значение имеет дофамин, выполняющий роль тормозного медиатора, а также медиатор ацетилхолин.
Бόльшая часть афферентных импульсов, приходящих к подкорковым ядрам, поступает в полосатое тело. Эти импульсы исходят почти всегда из трёх источников: 1) от всех областей коры больших полушарий; 2) от специфических ядер таламуса; 3) от чёрной субстанции среднего мозга по дофаминергическому пути.
Эфферентные пути стриопаллидарной системы главным образом выходят через бледный шар. Один путь образован аксонами расположенных в бледном шаре нейронов и идёт к различным ядрам промежуточного и среднего мозга, в том числе и к красным ядрам среднего мозга, от которых начинается руброспинальный экстрапирамидный путь, осуществ-
ляющий регуляцию движений. Второй важный эфферентный путь идёт от внутреннего отдела бледного шара к ядрам таламуса и далее к среднему мозгу. Таким образом, базальные ганглии играют главным образом роль промежуточного звена в цепи, связывающей двигательные области коры со всеми остальными её областями.
 Подкорковые ядра играют важную роль в регуляции движений и сенсомоторной координации. При поражении базальных ганглиев развиваются три типа серьёзных нарушений: 1) изменение мышечного тонуса, 2) утрата некоторых определённых движений, 3) появление нескольких видов непроизвольных движений.
Подкорковые ядра играют важную роль в регуляции движений и сенсомоторной координации. При поражении базальных ганглиев развиваются три типа серьёзных нарушений: 1) изменение мышечного тонуса, 2) утрата некоторых определённых движений, 3) появление нескольких видов непроизвольных движений.
Изменение мышечного тонуса при большинстве заболеваний подкорковых ядер заключается гла-вным образом в его усилении. Возникает гипертонус, или восковидная ригидность, не зависящая от положения суставов и фазы движения. Ригидность может иногда захватывать все группы мышц, которые бывают настолько напряжены, что выполнение произвольных движений практически невозможно. Чаще всего повышенный тонус развивается в мышцах, противодействующих силе гравитации. При пассивных движениях мышцы расслабляются не постепенно, а ступенчато («симптом зубчатого колеса»).
Утрата некоторых определённых движений, или акинезия, имеет различное выражение в зависимости от места и степени патологических изменений в подкорковых ядрах. Акинезия проявляется в скованности движений, неподвижной позе и маскообразном лице с немигающим выражением глаз. Больные испытывают большие, а иногда непреодолимые трудности при начале или завершении движения. Утрачиваются содружественные движения, обычно связанные с определёнными видами двига-
тельной активности. Например, при ходьбе отсутствуют содружественные движения рук. Больной встаёт и садится, не производя при этом целого ряда естественных содружественных движений туловища и конечностей.
Непроизвольные движения, возникающие при заболеваниях базальных ганглиев, включают разные виды двигательных расстройств. Среди них преобладают несколько видов дрожания, или тремора, хорея, атетоз и болезнь Паркинсона. Наиболее часто тремор проявляется в концах конечностей, однако может захватывать голову, лицо и туловище. Тремор, возникающий во время покоя, называется статическим тремором. Он подавляется при выполнении произвольных движений. Наиболее тяжёлую форму тремор приобретает при эмоциональном возбуждении.
Хорея – это заболевание, при котором происходят непроизвольные быстрые, иногда причудливые судорожные подёргивания, вовлекающие различные группы мышц. Это может быть сгибание пальца, отведение рук, подъём плеча, гримаса лица и т.п. Иногда эти движения кажутся целесообразными, однако, чаще производят впечатление бессмысленных действий. Они могут возникать не только в покое, но и сопровождать произвольные акты. Хорея, как правило, вызвана поражением полосатого тела.
Атетоз возникает, как и хорея, при поражении полосатого тела. Он проявляется в виде медленных червеобразных движений кистей и пальцев рук. Обычно наблюдаются чередующиеся сильные сгибания и разгибания кистей и пальцев рук. В тяжёлых случаях вовлекается лицевая мускулатура. Гримаса на лице часто возникает одновременно с движением конечности.
Болезнь Паркинсона, или дрожательный паралич, возникает при обширных поражениях нейронов подкорковых ядер и особенно бледного шара. При этой болезни также повреждаются чёрная субстанция среднего мозга и дофаминергический афферентный путь, идущий от чёрной субстанции к полосатому телу и, по-видимому, оказывающий на него тормозное влияние. Болезнь Паркинсона проявляется в виде комбинации тремора, ригидности и акинезии.
В последнее время для выяснения роли подкорковых ядер в осуществлении движений используют в опытах на животных вживленные микроэлектроды. В опытах на обезьянах установлено наличие взаимосвязи между разрядами нейронов полосатого тела и медленными, направленными из стороны в сторону червеобразными движениями конечностей. По современным представлениям, базальные ганглии являются одним из уровней иерархической системы регуляции движений. Получая информацию от ассоциативных зон коры, подкорковые ядра участвуют в создании программы целенаправленных движений. От подкорковых ядер далее информация поступает в передний таламус, где она объединяется с информацией, приходящей от мозжечка. Из ядер таламуса импульсы направля ются в двигательную кору, которая отвечает за осуществление программы целенаправленного движения через нижележащие двигательные центры, расположенные в стволе мозга и в спинном мозге.
Строение коры больших полушарий
Кора больших полушарий в филогенетическом плане является наиболее молодым образованием головного мозга. У рыб имеются только зачатки коры, у земноводных, рептилий и птиц размеры её постепенно увеличиваются. Наибольшего развития кора достигает у млекопитающих и особенно у человека. В соответствии с историческим процессом развития кору больших полушарий подразделяют на древнюю кору (архикортекс), старую кору (палеокортекс) и новую кору (неокортекс). К древней относят участок коры, называемый обонятельными луковицами, в которые поступают нервные волокна от органов обоняния, а также обонятельные проводящие пути, расположенные на нижней поверхности лобной доли, и обонятельные бугорки. Старая кора включает поясную извилину и гиппокамп. Все остальные области относятся к новой коре, которая у человека занимает 95,6 % всей площади коры.
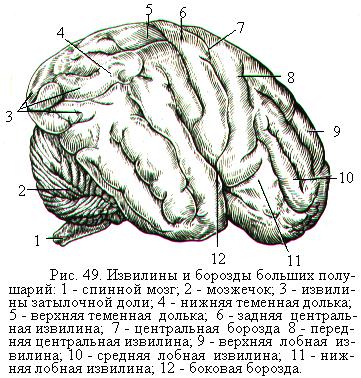 Поверхность больших полушарий покрыта тонким слоем (от 1,5 до 5 мм) серого вещества – плащом, или корой больших полушарий. Сверху на коре имеются многочисленные неровные углубления – борозды, между которыми выступают извилины (рис. 49). Борозды и извилины увеличивают поверхность коры до 1500 - 2000 см2 без увеличения объёма черепа. У человека около 65 % поверхности всей коры расположено в глубине борозд. Величина и форма борозд и извилин у каждого человека имеют значительные индивидуальные колебания. Однако существует несколько постоянных борозд, которые ясно выражены у всех и раньше других появляются в процессе развития зародыша. Эти борозды разделяют каждое из полушарий на большие участки, называемые долями.
Поверхность больших полушарий покрыта тонким слоем (от 1,5 до 5 мм) серого вещества – плащом, или корой больших полушарий. Сверху на коре имеются многочисленные неровные углубления – борозды, между которыми выступают извилины (рис. 49). Борозды и извилины увеличивают поверхность коры до 1500 - 2000 см2 без увеличения объёма черепа. У человека около 65 % поверхности всей коры расположено в глубине борозд. Величина и форма борозд и извилин у каждого человека имеют значительные индивидуальные колебания. Однако существует несколько постоянных борозд, которые ясно выражены у всех и раньше других появляются в процессе развития зародыша. Эти борозды разделяют каждое из полушарий на большие участки, называемые долями.
Центральная борозда отделяет лобную долю от теменной, боковая (латеральная, Сильвиева) борозда – височную долю от лобной и теменной, теменно-затылочная борозда разделяет теменную и затылочную до-
ли. В глубине боковой борозды располагается островковая доля. Иногда в качестве отдельной доли выделяют лимбическую долю, расположенную на внутренней поверхности большого полушария и включающую две извилины: поясную и парагиппокампову (рис. 50). Более мелкие борозды делят доли на извилины.
 Кора больших полушарий состоит из 10 – 18 млрд. нервных клеток, их отростков и нейроглии. Клетки нейроглии выполняют ряд важных функций: они являются опорной тканью, участвуют в обмене веществ головного мозга, регулируют кровоток внутри мозга, выполняют защитную функцию.
Кора больших полушарий состоит из 10 – 18 млрд. нервных клеток, их отростков и нейроглии. Клетки нейроглии выполняют ряд важных функций: они являются опорной тканью, участвуют в обмене веществ головного мозга, регулируют кровоток внутри мозга, выполняют защитную функцию.
Нейроны коры по своим размерам, форме и функциям подразделяются на несколько групп. Одна группа состоит из разных по размерам пирамидных клеток. Они расположены перпендикулярно по отношению к поверхности коры и имеют тело треугольной формы. От тела пирамидного нейрона вверх отходит Т-образно ветвящийся дендрит, а вниз от основания нейрона отходит аксон, который либо направляется к другим зонам коры, либо покидает кору в составе нисходящих нервных путей. На разветвлениях дендрита находится огромное количество мелких выростов – шипиков, являющихся областями синаптических контактов. Пирамидные нейроны делятся на крупные и мелкие. От крупных начинаются эфферентные пути к подкорковым структурам и далее к мотонейронам спинного мозга. Мелкие пирамидные нейроны образуют ассоциативные пути к другим участкам коры.
Другую группу нейронов коры составляют более мелкие звёздчатые клетки (клетки-зёрна). Они имеют сильно ветвящиеся дендриты и аксоны, образующие внутрикорковые связи.
Третья группа нейронов коры представлена веретеновидными клетками, имеющими длинный аксон, который отходит от тела клетки или вертикально или горизонтально.
Аксоны, отходящие от нейронов коры, образуют три разновидности нервных волокон: 1) проекционные волокна, по которым нервные им-
пульсы поступают к нижележащим структурам головного мозга и к спинному мозгу; 2) ассоциативные волокна, идущие к другим участкам коры того же полушария; 3) комиссуральные волокна, соединяющие области коры правого и левого полушария и образующие мозолистое тело.
 Величина, строение и взаиморасположение нейронов неодинаково в различных участках коры и на разной её глубине, что определяет архитектуру коры, или её цитоархитектонику. В связи с тем, что тела и отростки нейронов расположены упорядоченно, на по-перечном срезе коры выделяют 6 горизонтальных слоёв (рис. 51). Первый слой – это самый наружный молекулярный слой, содержащий незначительное число тел нейронов. Он состоит из густого сплетения нервных волокон, расположенных параллельно поверхности корковых извилин. Основную массу этих волокон составляют дендриты пирамидных нейронов, тела которых расположены в нижележащих слоях.
Величина, строение и взаиморасположение нейронов неодинаково в различных участках коры и на разной её глубине, что определяет архитектуру коры, или её цитоархитектонику. В связи с тем, что тела и отростки нейронов расположены упорядоченно, на по-перечном срезе коры выделяют 6 горизонтальных слоёв (рис. 51). Первый слой – это самый наружный молекулярный слой, содержащий незначительное число тел нейронов. Он состоит из густого сплетения нервных волокон, расположенных параллельно поверхности корковых извилин. Основную массу этих волокон составляют дендриты пирамидных нейронов, тела которых расположены в нижележащих слоях.
Во втором слое, называемом наружным зернистым, находится множество мелких звёздчатых нейронов. Встречаются в нём и малые пирамидные клетки.
Третий – самый широкий, наружный пирамидный слой содержит средней величины нейроны пирамидной формы.
Четвёртый слой – внутренний зернистый – состоит из множества звёздчатых клеток (клеток-зёрен), обусловливающих его зернистую структуру.
Пятый слой – внутренний пирамидный – образован крупными пирамидными клетками, которые были открыты В.А. Бецем и впоследствии названы гигантскими клетками Беца. Аксоны этих нейронов образуют
нисходящие нервные пути к подкорковым ядрам, продолговатому и спинному мозгу. По этим путям от корковых центров двигательной активности поступают импульсы, координирующие целенаправленные двигательные акты и позу тела.
 Шестой слой, называемый полиморфным, находится непосредственно над белым веществом больших полушарий. Он состоит из неодинаковых по величине клеток треугольной и веретенообразной формы. Аксоны этих нейронов формируют нисходящие пути к зрительным буграм промежуточного мозга.
Шестой слой, называемый полиморфным, находится непосредственно над белым веществом больших полушарий. Он состоит из неодинаковых по величине клеток треугольной и веретенообразной формы. Аксоны этих нейронов формируют нисходящие пути к зрительным буграм промежуточного мозга.
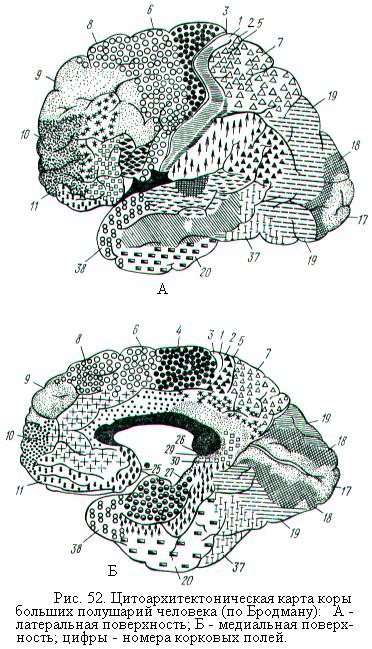
Выраженность отдельных слоёв в различных областях коры не одинакова. Учитывая эту особенность, К. Бродман по плотности расположения и форме нейронов разделил всю кору примерно на 52 цитоархитектонических поля (рис. 52). Впоследствии было установлено, что цитоархитектоническое разделение коры на поля в известной степени соответствует делению коры на зоны по функциональным и нейрохимическим особенностям. Поля, имеющие одинаковую функциональную специализацию, объединяют в области. Всего на карте коры больших полушарий человека выделяют 11 областей.
При изучении функций коры больших полушарий современными микроэлектродными методами было установлено, что кора состоит из элементарных функциональных единиц – колонок, расположенных перпендикулярно поверхности коры (колончатая организация коры). Каждая колонка включает в себя несколько структурных единиц - микромодулей, состоящих из пирамидных, звёздчатых и веретенообразных клеток, а так-
же волокон, кровеносных сосудов и клеток нейроглии. Микромодуль имеет диаметр около 100-150 мкм и объединяет 5-6 гнездообразно расположенных нейронов. Чаще всего это вытянутые по вертикали пирамидные клетки, с которыми контактируют звёздчатые нейроны. На звёздчатых клетках образуют синапсы нервные волокна, идущие от таламуса. Некоторые из звёздчатых нейронов, выполняющие тормозную функцию, имеют длинные аксоны, идущие в горизонтальном направлении.
Нейроны одной колонки отвечают на раздражитель только одной разновидности и являются элементарным блоком, где происходит локальная переработка информации от рецепторов. Обычно возбуждение одного из модулей сопровождается торможением соседних, что препятствует распространению возбуждения и, таким образом, усиливает контрастное различение контуров раздражения.
Функциональное значение коры больших полушарий
Для изучения функций коры больших полушарий применяют различные методы: 1) удаление отдельных участков коры; 2) метод раздражения с использованием вживленных микроэлектродов, химических и температурных раздражителей; 3) метод отведения биопотенциалов от отдельных зон и нейронов коры больших полушарий; 4) метод условных рефлексов, разработанный И.П. Павловым; 5) клинический метод, позволяющий изучать деятельность отдельных органов и их систем, которые наблюдаются у людей при повреждении коры больших полушарий.
С появлением в процессе эволюции коры больших полушарий она начинает контролировать все процессы, протекающие в организме, а также всю деятельность человека, т.е. происходит кортиколизация функций. Кора больших полушарий выполняет следующие функции: 1) осуществляет высшую регуляцию всех двигательных и эмоционально-вегетативных реакций; 2) производит высший анализ и синтез всей информации, поступающей от рецепторов; 3) обеспечивает замыкательную функцию, т.е. образование новых условных рефлексов и их систем – динамических стереотипов; 4) благодаря памяти в коре накапливается огромный объём информации; 5) за счёт деятельности коры осуществляются высшие психические функции человека: мышление, сознание, речь.
По выполнению функций обнаружена определённая специализация различных областей коры, т.е. выявлена локализация функций в коре больших полушарий. С этой точки зрения древнюю и старую кору выделили в особую систему конечного мозга, названную обонятельным, или висцеральным, мозгом, потому что эти области коры осуществляют функции, связанные с обонянием, управляют реакциями настораживания и внимания, принимают участие в регуляции вегетативных функций. Висцеральный мозг играет также важную роль в осуществлении инстинктивного поведения (пищевого, полового, оборонительного) и в формировании эмоций.
Раздражение структур старой коры оказывает влияние на сердечно-сосудистую систему и дыхание. Повреждения в области старой коры могут вызвать гиперсексуальность, изменять эмоциональное поведение. Существует тесная связь структур старой коры с вегетативной нервной системой, с процессами, направленными на регуляцию внутренней среды организма. Эта деятельность старой коры протекает при участии гипоталамуса.
Некоторые области старой коры, в частности гиппокамп, имеют важное значение в процессах памяти. У больных с нарушениями памяти при патоморфологических исследованиях часто обнаруживаются патологические изменения в гиппокампе.
Использование методов удаления и электрофизиологического исследования новой коры позволило обнаружить в ней две группы полей, или зон: проекционные, которые в свою очередь подразделяются на первичные проекционные и вторичные проекционные, и ассоциативные. Первичные проекционные зоны (первичные поля) связаны с органами чувств и органами движения. Они являются центральными, или корковыми, отделами анализаторов (зрительная зона, слуховая зона и т.д.). Первичные зоны получают импульсы от одинаковых рецепторов (одной модальности), но не могут полностью обеспечить их анализ и интегративное восприятие. При разрушении первичных проекционных зон возникает так называемая корковая слепота, корковая глухота и т.д.
Рядом с первичными проекционными зонами расположены вторичные проекционные зоны (вторичные поля), которые связаны с органами тела через первичные зоны. Вторичные проекционные зоны служат для обобщения и дальнейшей обработки информации, поступающей от определённых рецепторов. При их разрушении человек видит предметы, но не узнаёт их, или слышит слова, но не понимает их значения.
В коре больших полушарий человека обнаружены следующие проекционные зоны: мотосенсорные, сенсомоторные, зрительные, слуховая, вкусовая, обонятельная и вестибулярная,
В 1870 г. Г. Фрич и Е. Гитциг обнаружили, что у собак и кошек электрическая стимуляция некоторых областей коры больших полушарий вызывает сокращение мышц противоположной стороны тела. Вскоре были определены границы двигательной области (моторной зоны) коры, которая у человека занимает переднюю центральную (прецентральную) извилину (рис. 53). В 30-е годы 20-го века У. Пенфилд, основываясь на результатах, полученных во время нейрохирургических операциях на человеке, установил точное представительство мышц тела и конечностей в передней центральной извилине. Слабое электрическое раздражение отдельных участков передней центральной извилины вызывало сокращение строго определённых мышц. В верхних частях этой извилины проецируется мускулатура ног, в средних – рук и большинства мышц туловища, в нижних – мышц лица и головы. Площадь участка коры в передней центральной извилине, связанного с какой-то мышцей, пропорциональна не размерам этой мышцы, а точности её движений. Наибольшее пространство в двигательной области коры занимает представительство мышц лица, языка, кисти руки.
Впоследствии на медиальной поверхности полушария рядом с передней центральной извилиной была обнаружена ещё одна вторичная двигательная область, при раздражении которой возникают сокращения мышц нижней части туловища. Обе двигательные области не только посылают импульсы к мышцам тела, но также получают информацию от кожных и мышечных рецепторов, т.е. имеют самостоятельные сенсорные входы. Поэтому они получили название первичной и вторичной мотосенсорной зоны. При повреждении этих зон наступают различные параличи (утрата движений) или парезы (ослабление движений).
Сенсорные импульсы от рецепторов кожи, мышц и внутренних органов поступают в заднюю центральную (постцентральную) извилину, являющуюся первой соматосенсорной зоной (рис. 53). В районе боковой борозды, отделяющей височную долю от теменной и лобной, обнаружена вторая соматосенсорная зона, которая имеет значительно меньшую площадь, чем первая. Как и в мотосенсорных зонах, в соматосенсорных зонах наибольшую площадь занимает представительство мышц лица, языка и кисти руки. При раздражении определённых точек соматосенсорной зоны одного полушария на противоположной половине тела в соответствующих участках возникали ощущения прикосновения, давления, покалывания, онемения, тепла или холода. Повреждение участков соматосенсорной зоны приводит к сильному ослаблению чувствительности той части тела, которая представлена в повреждённом участке коры. Первая и вторая соматосенсорные зоны кроме сенсорных входов содержит моторные выходы, и поэтому их часто называют первичной и вторичной сенсомоторными зонами.
Мотосенсорные и сенсомоторные зоны функционально взаимосвязаны, однако, в восприятии кожно-мышечной чувствительности главную роль играет первичная сенсомоторная зона, а в управлении движениями – первичная мотосенсорная зона.
Из числа других проекционных зон коры чёткую локализацию в затылочной доле в области шпорной борозды имеет первичная зрительная зона (рис. 53). В связи с бинокулярным зрением первичная зрительная зона каждого полушария получает зрительную информацию от сетчаток обоих глаз. Причём в каждое полушарие проецируются половины сетчаток (левые половины – в правое полушарие, а правые – в левое). При этом в каждом полушарии происходит совмещение зрительных полей обоих глаз, что обеспечивает восприятие объёма предметов и других эффектов бинокулярного зрения. Электрическое раздражение сенсорной зрительной зоны может вызвать простейшие зрительные ощущения. Рядом с первичной расположена вторичная зрительная зона, участвующая в обеспечении зрительного внимания и управлении движениями глаз.
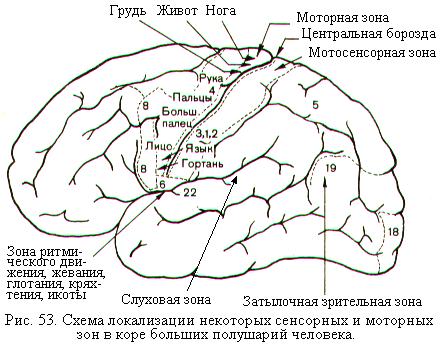 Сенсорная слуховая зона расположена в височной области в основном в глубине боковой борозды (рис. 53). Только небольшая часть этой зоны видна на верхнем крае височной доли. Различные участки кортиева органа улитки проецируются в определённые участки слуховой зоны. Электрическое раздражение слуховой зоны у людей с сохранённым сознанием вызывает ощущения шума в обоих ушах. Благодаря наличию слуховой зоны в обоих полушариях повреждение слуховой зоны в одном полушарии затрудняет восприятие речи, определение источника звука в пространстве и распознавание временных характеристик звука, но не приводит к глухоте. Удаление обеих слуховых зон вызывает глухоту.
Сенсорная слуховая зона расположена в височной области в основном в глубине боковой борозды (рис. 53). Только небольшая часть этой зоны видна на верхнем крае височной доли. Различные участки кортиева органа улитки проецируются в определённые участки слуховой зоны. Электрическое раздражение слуховой зоны у людей с сохранённым сознанием вызывает ощущения шума в обоих ушах. Благодаря наличию слуховой зоны в обоих полушариях повреждение слуховой зоны в одном полушарии затрудняет восприятие речи, определение источника звука в пространстве и распознавание временных характеристик звука, но не приводит к глухоте. Удаление обеих слуховых зон вызывает глухоту.
Вкусовая зона располагается в теменной доле, в нижней части задней центральной извилины. При её раздражении возникают различные вкусовые ощущения.
Обонятельная зона коры располагается на основании мозга, в области гиппокамповой извилины. В возникновении обонятельных ощущений большое значение имеют структуры древней коры.
Местонахождение вестибулярной зоны у человека окончательно не выяснено. У обезьян вестибулярная зона расположена впереди от нижнего участка центральной борозды.
Первичные и вторичные проекционные зоны (первичные и вторичные поля) хорошо выражены как у человека, так и у позвоночных животных. В коре больших полушарий человека и млекопитающих рядом с проекционными зонами расположены поля, которые не связаны с выполнением какой-либо специфической сенсорной или моторной функции. Такие поля составляют ассоциативные зоны (третичные поля). Они либо отсутствуют, либо слабо развиты даже у млекопитающих животных.
Только у обезьян они сравнительно хорошо представлены, а у человека занимают более 1/3 площади коры.
Нейроны ассоциативных зон реагируют на раздражения различных модальностей, причём их ответы возникают не на отдельные элементы объекта, а на целые его комплексы. Ассоциативные зоны участвуют в интеграции сенсорной информации и в обеспечении связей между чувствительными и двигательными зонами коры. Эти механизмы являются физиологической основой высших психических функций. Получив максимальное развитие у человека, ассоциативные зоны участвуют в осуществлении новых, специфически человеческих функций: речи, чтения, письма, логического мышления, интеллекта и т.д.
Ассоциативные зоны занимают ряд областей теменной, височной и лобной долей коры. В настоящее время установлено, что теменные ассоциативные зоны участвуют в оценке биологически значимой информации и в восприятии расположения окружающих предметов в пространстве. Височные ассоциативные зоны обеспечивают понимание речи. Бόльшую часть лобных долей составляют ассоциативные зоны, которые имеют обширные двусторонние связи с лимбической системой. Лобные ассоциативные зоны контролируют оценку мотивации поведения и программирование сложных поведенческих актов, участвуют в управлении движениями. Так, например, поражение лобных долей вызывает у больных тенденцию к повторению двигательных актов без видимой на то необходимости, а также нарушает запоминание месторасположения предметов. Предполагается, что лобные ассоциативные зоны наряду с теменными участвуют в интеграции сведений о времени и пространстве.
Таким образом, к настоящему времени накоплен большой экспериментальный материал, указывающий на локализацию функций преимущественно в каких-то определённых участках коры больших полушарий. В то же время получены новые данные, свидетельствующие о том, что управление какими-либо навыками не находится в каком-то одном центре. Даже при таком простом поведенческом акте, как взятие предмета пальцами, координация движений руки и пальцев осуществляется с помощью зрения. Очевидно, что для выполнения этого акта необходимы связи между зрительной зоной коры, расположенной в затылочной доле, и центрами управления движениями, т.е. мотосенсорной зоной в лобной доле и сенсомоторной зоной в теменной доле. Кроме того, для зрительного слежения за пальцами необходима произвольная регуляция движений глаз, поэтому в данном акте должны участвовать центры управления глазами в лобных долях. Анатомические исследования показывают, что между всеми этими зонами имеются нервные связи.
Становится ясно, что даже в простом поведенческом акте должны активно участвовать обширные зоны коры, расположенные в различных долях и даже полушариях. Тем более нельзя считать, что управление высшими психическими функциями является компетенцией какого-то одного центра. Таким образом, сегодня на смену старому представлению о локализации функций в отдельных зонах коры приходит новая концепция, согласно которой структурную основу мозговых функций составляют распределённые системы. Каждую функцию обслуживает распределённая система, состоящая из определённого сочетания корковых и подкорковых центров, соединённых между собой прямыми и обратными нервными связями.
Электрическая активность коры больших полушарий
Для изучения функциональной активности коры больших полушарий и её взаимоотношений с подкорковыми структурами используется метод регистрации биотоков мозга – электроэнцефалография (ЭЭГ). Сама запись отведённых с поверхности кожи головы и усиленных в сотни тысяч раз этих электрических колебаний на бумаге или фотоплёнке называется электроэнцефалограммой. При нейрохирургических операциях на человеке ЭЭГ регистрируют непосредственно от поверхности коры больших полушарий. В этом случае её называют электрокортикограммой (ЭКоГ). По сравнению с ЭЭГ электрокортикограмма имеет более высокую амплитуду и частоту быстрых колебаний.
ЭЭГ представляет собой результат алгебраического сложения синаптических потенциалов, возникающих в расположенных у поверхности коры множестве дендритов корковых пирамидных нейронов. Причиной синхронного возбуждения нейронов коры и возникновения одновременных изменений постсинаптических потенциалов могут быть импульсы, поступающие от ядер таламуса. В ядрах таламуса обнаружена ритмическая электрическая активность, частота которой совпадает с альфа-ритмом ЭЭГ. Эта ритмическая активность может быть обусловлена либо свойствами самих клеток таламуса, либо активностью нейронных сетей с возбуждающими и тормозными синаптическими связями. Поступление сенсорных импульсов в кору (например, при открывании глаз, при умственной деятельности) приводит к нарушению синхронности колебаний ЭЭГ.
При анализе ЭЭГ учитывают частоту, амплитуду, форму и длительность электрических колебаний. В настоящее время различные параметры ЭЭГ достаточно хорошо изучены и анализ изменений ЭЭГ широко используется при оценке состояния различных областей коры и в клинической практике.
В ЭЭГ взрослого человека выделяют 4 основных типа ритмических электрических колебаний (ритмов ЭЭГ), каждый из которых характерен для определённого функционального состояния (рис. 54).
Альфа-ритм (α), с частотой 8-13 колебаний в 1 секунду и амплитудой примерно 50 мкВ, регистрируется в состоянии спокойного бодрствования и при отсутствии внешних раздражений. У человека альфа-ритм наиболее выражен в затылочных долях коры и хорошо регистрируется, когда он сидит спокойно с закрытыми глазами. Как только человек открывает глаза, альфа-ритм сменяется бета-ритмом.
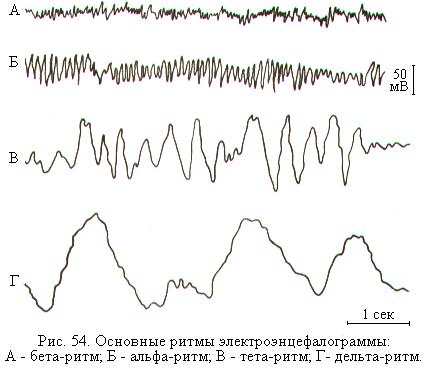 Бета-ритм (β), с частотой 14-30 колебаний в 1 секунду и амплитудой примерно 25 мкВ, характерен для активного бодрствования. Этот ритм наблюдается преимущественно в лобных и теменных областях коры больших полушарий. Переход человека от покоя к состоянию активного внимания, к умственной деятельности, а также восприятие зрительных или слуховых сигналов сопровождается исчезновением альфа-ритма и возникновением колебаний бета-ритма. Это получило название реакции десинхронизации, или реакции активации. Противоположностью десинхронизации является реакция синхронизации или инактивации. При синхронизации уровень возбуждения в коре снижается и вместо бета-ритма на ЭЭГ появляется альфа-ритм.
Бета-ритм (β), с частотой 14-30 колебаний в 1 секунду и амплитудой примерно 25 мкВ, характерен для активного бодрствования. Этот ритм наблюдается преимущественно в лобных и теменных областях коры больших полушарий. Переход человека от покоя к состоянию активного внимания, к умственной деятельности, а также восприятие зрительных или слуховых сигналов сопровождается исчезновением альфа-ритма и возникновением колебаний бета-ритма. Это получило название реакции десинхронизации, или реакции активации. Противоположностью десинхронизации является реакция синхронизации или инактивации. При синхронизации уровень возбуждения в коре снижается и вместо бета-ритма на ЭЭГ появляется альфа-ритм.
|
|
|
|
|
Дата добавления: 2014-12-26; Просмотров: 1408; Нарушение авторских прав?; Мы поможем в написании вашей работы!