КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Строение синапса. Синаптическая передача возбуждения ….….. 23 5 страница
|
|
|
|
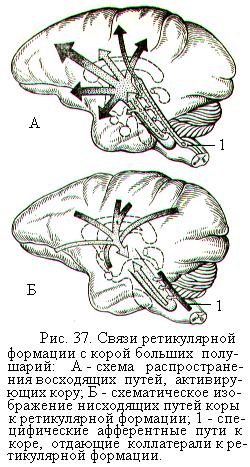 Активность самой ретикулярной формации поддерживается непрерывным поступлением импульсов, идущих от рецепторов тела. Важная роль в поддержании её активности принадлежит гуморальным факторам, по отношению к которым она обладает высокой чувствительностью.
Активность самой ретикулярной формации поддерживается непрерывным поступлением импульсов, идущих от рецепторов тела. Важная роль в поддержании её активности принадлежит гуморальным факторам, по отношению к которым она обладает высокой чувствительностью.
Благодаря работам Х. Мегуна и Дж. Моруцци были открыты восходящие, активирующие влияния ретикулярной формации на кору больших полушарий (рис. 37). Обнаружено, что ретикулярная формация участвует в регуляции сна и бодрствования. Раздражение и возбуждение её с помощью вживленных в мозг электродов вызывает пробуждение у спящих животных. Эта поведенческая реакция пробуждения сопровождается учащением ритма электроэнцефалограммы в обширных областях коры больших полушарий. Разрушение восходящих путей от ретикулярной формации приводит к глубокому сну у бодрствовавших животных и уменьшению частоты колебаний электроэнцефалограммы.
Большинство нейронов ретикулярной формации являются полисенсорными, т.е. отвечают на различные раздражения: световые, звуковые, тактильные и т.д. Эти нейроны имеют обширные рецептивные поля, большой латентный период и слабую воспроизводимость реакции, что сильно отличает их от нейронов специфических ядер. В связи с этим нейроны ретикулярной формации относят к неспецифическим. Точно также восходящие пути ретикулярной формации называют неспецифическими, т.к. они направлены к обширным областям коры больших полушарий в
отличие от специфических проекционных путей от органов чувств, идущих в конкретные зоны коры.
Ретикулярной формации принадлежит важная роль в механизмах формирования условнорефлекторных реакций организма. Она повышает активность вегетативных нервных центров, функционируя совместно с симпатическим отделом вегетативной нервной системы. Введение адреналина повышает тонус ретикулярной формации, в результате чего усиливается её активирующее влияние на кору больших полушарий. Адреналин, выделяемый мозговым веществом надпочечников при эмоциях, действуя на ретикулярную формацию, увеличивает и удлиняет эффекты возбуждения симпатической нервной системы.
Благодаря наличию кольцевых связей в ретикулярной формации происходит взаимодействие афферентных и эфферентных импульсов, возможна их продолжительная циркуляция по кругу. Вследствие этого поддерживается определённый уровень возбуждения самой ретикулярной формации, а она в свою очередь поддерживает тонус и готовность к деятельности различных отделов центральной нервной системы. Активность ретикулярной формации находится под регулирующим влиянием коры больших полушарий.
МОЗЖЕЧОК
Строение мозжечка
Мозжечок расположен позади и чуть выше продолговатого мозга и варолиева моста под большими полушариями. Это надсегментарная структура, появляющаяся на ранних этапах филогенеза хордовых животных. Степень развития мозжечка определяется сложностью среды обитания и передвижения организма. Наибольшего развития мозжечок достигает у человека в связи с прямохождением и усложнением движений при трудовой деятельности. В то же время мозжечок не является жизненно необходимым органом. У людей с врождённым отсутствием мозжечка не наблюдается каких-либо серьёзных нарушений движений, препятствующих их жизнедеятельности.
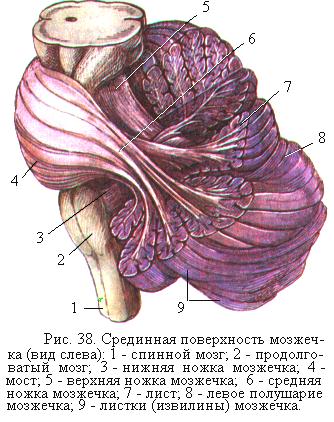
Мозжечок человека состоит из непарной средней части – червя и расположенных по обе стороны от червя двух полушарий. Со стволом мозга мозжечок соединяется тремя парами ножек. Толстые средние ножки охватывают продолговатый мозг и, расширяясь, переходят в варолиев мост (рис. 38). Верхние ножки мозжечка идут к четверохолмию среднего мозга, а нижние ножки спускаются вниз и сливаются с продолговатым мозгом. Через нижние и средние ножки главным образом входят в мозжечок афферентные волокна, а через верхние ножки выходят эфферентные волокна.
Поверхность мозжечка покрыта серым веществом, толщиной 1 – 2,5 мм, образующим его кору. Под корой находится белое вещество, в котором группами располагается серое вещество, представляющее собой скопления тел нейронов – ядра мозжечка. На поверхности полушарий и червя имеются поперечные параллельные борозды, между которыми расположены узкие длинные листки мозжечка. Каждый листок (извилина) мозжечка представляет собой тонкую прослойку белого вещества, покрытого корой. Листки группируются в дольки, которые отделяются более глубокими бороздами. Складчатое строение во много раз увеличивает поверхность коры мозжечка (рис. 38).
 На поперечном срезе коры мозжечка различают три слоя: наружный – молекулярный, средний – ганглиозный и внутренний – зернистый (рис. 39). Молекулярный слой составляют: 1) параллельные волокна, представляющие собой Т-образные разветвления аксонов многочисленных клеток-зёрен, тела которых находятся во внутреннем слое; 2) разветвления дендритов клеток Пуркинье, клеток Гольджи, а также аксонов клеток-зёрен, тела которых залегают в нижележащих слоях; 3) корзинчатые клетки; 4) некоторое количество звёздчатых клеток.
На поперечном срезе коры мозжечка различают три слоя: наружный – молекулярный, средний – ганглиозный и внутренний – зернистый (рис. 39). Молекулярный слой составляют: 1) параллельные волокна, представляющие собой Т-образные разветвления аксонов многочисленных клеток-зёрен, тела которых находятся во внутреннем слое; 2) разветвления дендритов клеток Пуркинье, клеток Гольджи, а также аксонов клеток-зёрен, тела которых залегают в нижележащих слоях; 3) корзинчатые клетки; 4) некоторое количество звёздчатых клеток.
В ганглиозном слое находятся тела около 7 млн. клеток Пуркинье, ориентированные вертикально по отношению к поверхности коры мозжечка. Их дендриты поднимаются вверх в молекулярный слой, где сильно ветвятся. На дендритах клеток Пуркинье находится множество шипиков, на которых образуют синапсы параллельные волокна. Аксоны клеток Пуркинье выходят из коры мозжечка вниз и направляются к ядрам мозжечка. Аксоны клеток Пуркинье являются практически единственным выходом из коры мозжечка.
Под ганглиозным слоем лежит зернистый, или гранулярный, слой. В нём насчитывается свыше 10 млрд. клеток-зёрен, или гранулярных клеток. Аксоны этих клеток поднимаются вертикально вверх в молекулярный слой, где Т-образно ветвятся. Ветви идут параллельно поверхности коры (параллельные волокна) и образуют синапсы на дендритах других клеток. В зернистом слое находятся тела клеток Гольджи, дендриты которых поднимаются вверх и разветвляются в молекулярном слое, а аксоны образуют синапсы на клетках-зёрнах.
За исключением клеток-зёрен, все вышеописанные нервные клетки, тела которых расположены в коре мозжечка, являются тормозными.
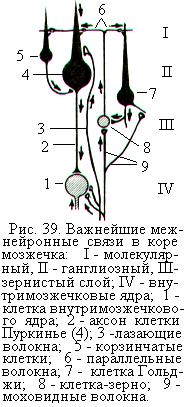 Вход в кору мозжечка осуществляется по трём типам афферентных волокон: моховидным, лиановидным и норадренергическим. Моховидные, или мшистые, волокна идут от нейронов ядер варолиева моста и образуют синапсы на клетках-зёрнах. Лиановидные, или лазающие, волокна идут из продолговатого мозга. Они сильно ветвятся, оплетая дендриты клеток Пуркинье и формируя на них многочисленные возбуждающие синапсы (рис. 39). Норадренергические волокна поступают из голубого пятна, которое расположено в боковой части ромбовидной ямки, или дна IV желудочка, образованного задними поверхностями продолговатого мозга и моста.
Вход в кору мозжечка осуществляется по трём типам афферентных волокон: моховидным, лиановидным и норадренергическим. Моховидные, или мшистые, волокна идут от нейронов ядер варолиева моста и образуют синапсы на клетках-зёрнах. Лиановидные, или лазающие, волокна идут из продолговатого мозга. Они сильно ветвятся, оплетая дендриты клеток Пуркинье и формируя на них многочисленные возбуждающие синапсы (рис. 39). Норадренергические волокна поступают из голубого пятна, которое расположено в боковой части ромбовидной ямки, или дна IV желудочка, образованного задними поверхностями продолговатого мозга и моста.
В белом веществе мозжечка человека находятся парные ядра: ядро шатра, вставочное, или промежуточное ядро, которое разделяется на шаровидное и пробковидное ядра, и зубчатое ядро. Эфферентные волокна разных участков коры мозжечка, являющиеся аксонами клеток Пуркинье, направляются к строго определённым ядрам мозжечка. В соответствии с проекцией коры мозжечка на его ядра кору мозжечка подразделяют на три продольные зоны – кору червя, кору промежуточной околочервячной зоны и кору остальной части полушарий мозжечка. Так, эфферентные волокна от коры червя мозжечка идут к ядру шатра, от промежуточной околочервячной зоны коры – к вставочному ядру, а от остальных боковых участков коры полушарий мозжечка – к зубчатому ядру.
Функции мозжечка
Мозжечок выполняет проводниковую, рефлекторную и интегративную функции. По афферентным спинно-мозжечковым путям в мозжечок поступают импульсы от рецепторов кожи, мышц и сухожилий. От вестибулярных ядер продолговатого мозга по вестибуло-мозжечковым путям в мозжечок поступает информация о положении тела. Кора больших полушарий также посылает афферентные пути в мозжечок, среди которых наиболее важными являются кортико-мосто-мозжечковый и кортико-ретикуло-мозжечковый пути.
Эфферентные пути от мозжечка идут к спинному и продолговатому мозгу, к ретикулярной формации, красным ядрам среднего мозга, к промежуточному мозгу, коре больших полушарий и к подкорковым ядрам.
Мозжечок оказывает рефлекторные влияния на различные двигательные и вегетативные функции. Главное его значение заключается в дополнении и коррекции деятельности остальных двигательных центров. Мозжечок участвует: 1) в регуляции позы и мышечного тонуса; 2) в коррекции медленных целенаправленных движений и координации их с рефлексами поддержания позы; 3) в координации быстрых целенаправленных движений, осуществляемых по команде из коры больших полушарий.
Зона коры червя мозжечка главным образом связана с регуляцией позы, равновесия и мышечного тонуса. Промежуточная околочервячная зона коры мозжечка принимает участие в координации медленных целенаправленных движений с рефлексами поддержания позы.
Боковые участки коры, расположенные на полушариях мозжечка, участвуют в осуществлении быстрых целенаправленных движений. К полушариям мозжечка от ассоциативных областей коры больших полушарий поступает информация о замысле движения по афферентному кортико-мосто-мозжечковому пути, насчитывающему около 20 млн. нервных волокон. В полушариях мозжечка и зубчатом ядре мозжечка информация о замысле движения преобразуется в программу движения, которая через таламус промежуточного мозга посылается в двигательные зоны коры больших полушарий. От коры больших полушарий по нисходящим путям передаются команды к мотонейронам спинного мозга и происходит осуществление движений.
Мозжечок вносит в выполнение движений необходимые поправки, обеспечивая точность, ловкость и координированность движений. При нарушении функций мозжечка возникают различные двигательные расстройства: атония, астения, астазия, атаксия, асинергия, дисметрия, адиадохокинез, дезэквилибрия (рис. 40).
 Атония характеризуется резким ослаблением мышечного тонуса. Она обычно сопровождается астенией – слабостью и повышенной утомляемостью мышц. Астазия проявляется в утрате способности мышц к длительному тетаническому сокращению, вследствие чего конечности и голова непрерывно дрожат и качаются, т.е. наблюдается тремор. Атаксия характеризуется нарушением координации движений, походки и др. При атаксии ноги широко расставлены, движения избыточны, из-за которых больного как пьяного бросает из стороны в сторону.
Атония характеризуется резким ослаблением мышечного тонуса. Она обычно сопровождается астенией – слабостью и повышенной утомляемостью мышц. Астазия проявляется в утрате способности мышц к длительному тетаническому сокращению, вследствие чего конечности и голова непрерывно дрожат и качаются, т.е. наблюдается тремор. Атаксия характеризуется нарушением координации движений, походки и др. При атаксии ноги широко расставлены, движения избыточны, из-за которых больного как пьяного бросает из стороны в сторону.
Асинергия проявляется в нарушении взаимодействия между двигательными центрами различных мышц. При этом происходит как бы рас-
пад программы движения и целостное движение состоит не из одновременных содружественных актов, а распадается на ряд простых движений выполняемых последовательно. Асинергия сочетается с дисметрией, или утратой соразмерности движения. При дисметрии наблюдается несоответствие между интенсивностью мышечного сокращения и задачей выполняемого движения, движения становятся размашистыми и неупорядоченными в пространстве.
Адиадохокинез характеризуется нарушением координации быстрых целенаправленных движений. При этом человек не способен выполнять быструю последовательность движений, например поочередное сгибание и разгибание пальцев. При нарушении функций мозжечка также наблюдается дезэквилибрия, т.е. потеря способности сохранять равновесие.
Мозжечок участвует не только в регуляции движений, но и осуществляет контроль над вегетативными функциями, оказывая облегчающее или угнетающее влияние на деятельность сердечно-сосудистой, дыхательной, пищеварительной систем, на терморегуляцию. Влияния мозжечка осуществляются, по-видимому, благодаря его связям с ретикулярной формацией и гипоталамусом. Деятельность мозжечка протекает в непосредственной связи с корой больших полушарий и под её контролем.
ПРОМЕЖУТОЧНЫЙ МОЗГ
Структуры промежуточного мозга. Функции таламуса
Промежуточный мозг расположен под мозолистым телом и сверху полностью покрыт большими полушариями. Промежуточный мозг и большие полушария часто объединяют под названием передний мозг. В промежуточном мозге различают парные зрительные бугры, или таламус, подбугорную область, или гипоталамус, забугорную область, или коленчатые тела, и надбугорную область. Внутри промежуточного мозга имеется полость – третий желудочек, имеющий вид вертикальной щели и сообщающийся сверху с боковыми желудочками больших полушарий, а снизу переходящий в водопровод мозга.
Надбугорная область относительно мала. В ней расположена железа внутренней секреции – эпифиз.
Забугорную область, или латеральное и медиальное коленчатые тела, по функциональному значению часто объединяют со специфическими ядрами таламуса. Латеральное коленчатое тело является специфическим ядром зрительной системы, имеющим прямые связи со зрительными проекционными зонами, находящимися в затылочной доле коры больших полушарий. Нейроны этого ядра принимают участие в анализе освещённости, контраста, яркости, цвета и другой зрительной информации, поступающей от рецепторов зрительного анализатора.
В медиальное коленчатое тело идут через нижние бугры четверохолмия восходящие пути, проводящие импульсы от рецепторов слухового анализатора. Это тело является специфическим ядром слуховой системы, принимающим участие в анализе и передаче акустической информации. От нейронов медиального коленчатого тела начинается тракт, направляющийся в слуховые проекционные зоны, расположенные в височной доле коры больших полушарий.
 Наиболее важными функциональными образованиями промежуточного мозга являются таламус и гипоталамус.
Наиболее важными функциональными образованиями промежуточного мозга являются таламус и гипоталамус.
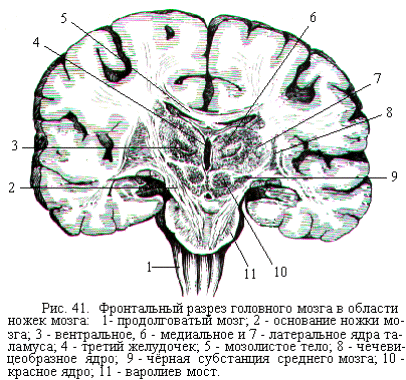
Таламус имеет форму двух крупных тел эллипсоидной формы, по бокам срастающихся с большими полушариями. Серое вещество в таламусе сгруппировано в ядра, которые расположены главным образом в области боковой стенки III-го желудочка (рис. 41). Всего в таламусе насчитывают до 40 ядер. По функциональному значению ядра подразделяют на специфичес-кие, неспецифические, ассоциативные и моторные.
На нейронах спе-цифических ядер заканчиваются волокна различных восходящих путей. В специфических, или проекционных, ядрах таламуса происходит синаптическая передача импульсов, поступивших от всех рецепторов тела, за исключением обонятельных, на нейроны, по аксонам которых импульсы направляются в соответствующие участки коры больших полушарий. Тот факт, что афферентные сигналы на пути к коре переключаются на нейронах таламуса, имеет большое значение. Тормозные влияния, приходящие в таламус из коры больших полушарий, других образований и соседних таламических ядер, позволяют подавить слабые второстепенные возбуждения и обеспечить лучшую передачу в кору наиболее важной информации.
Специфические ядра на пути от рецепторов к коре больших полушарий являются как бы передаточной станцией и одновременно фильтром, пропускающим к коре только наиболее важную информацию и тормозящим второстепенную. Нейроны специфических ядер посылают по направлению к коре аксоны, почти не имеющие боковых ответвлений. При одиночном раздражении каких-либо специфических ядер возникает
возбуждение в строго определённых точках коры, а при их повреждении пропадает определённый вид чувствительности.
Неспецифические ядра таламуса имеют многочисленные нервные связи с ретикулярной формацией, со специфическими и ассоциативными ядрами таламуса и со всеми областями коры больших полушарий. Нейроны неспецифических ядер посылают к коре аксоны, дающие множество боковых ответвлений. По ним сначала сигналы передаются в подкорковые структуры, от которых импульсы поступают параллельно в разные отделы коры. Через неспецифические ядра в кору передаются возбуждающие влияния от ретикулярной формации, в результате чего активность коры повышается. При раздражении неспецифических ядер возбуждение возникает и распространяется почти на все области коры. Реакция в коре возникает с большим скрытым периодом и значительно усиливается при повторении. Нейроны коры больших полушарий вовлекаются в процесс активности постепенно, что получило название реакции вовлечения.
Неспецифические ядра таламуса осуществляют контроль ритмической активности коры больших полушарий. Повреждения неспецифических ядер таламуса у людей при нейрохирургических операциях приводят к нарушениям сознания. Это свидетельствует о том, что импульсы, поступающие от неспецифических ядер, поддерживают уровень возбудимости корковых нейронов, необходимый для сохранения сознания.
Кора больших полушарий, в свою очередь, может оказывать тормозные и возбуждающие влияния на специфические и неспецифические ядра таламуса. Эти влияния распространяются на специфические ядра по прямым кортикоталамическим путям, а на неспецифические ядра через ретикулярную формацию. Например, раздражение сенсомоторной зоны коры угнетает на длительное время возникновение реакции вовлечения. Это свидетельствует о наличии между таламусом и корой больших полушарий двусторонних циклических связей, которые играют важную роль в интегративной деятельности мозга, в регуляции таких физиологически важных состояний, как сохранение сознания, смена сна и бодрствования.
Ассоциативные ядра таламуса не относятся к какой-либо одной сенсорной системе и получают афферентные импульсы только от специфических (проекционных) ядер. Через ассоциативные ядра таламус связан с ассоциативными областями коры.
Моторные ядра таламуса связаны с мозжечком, подкорковыми узлами, моторной зоной коры больших полушарий и участвуют в регуляции движений.
Функции гипоталамуса
Гипоталамус расположен под таламусом и кпереди от ножек мозга. Он включает в себя такие анатомические структуры, как серый бугор, воронку и сосцевидные тела (рис. 42). Гипоталамус имеет тесные нервные и сосудистые связи с рядом расположенной эндокринной железой – гипофизом, а также многочисленные нервные связи со всеми отделами ЦНС. Гипоталамус играет важную роль в поддержании постоянства внутренней среды организма, обеспечении интеграции функций соматической, вегетативной и эндокринной систем, а также в организации инстинктивного поведения.
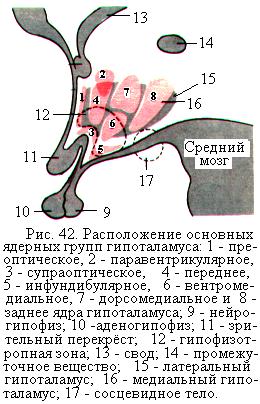 В гипоталамусе насчитывается более 30 пар ядер, которые на основе цитоархитектонического строения подразделяют на три не резко разграниченные группы: переднюю, среднюю и заднюю. С функциональной точки зрения ядра гипоталамуса делят на 5 групп: преоптическую, переднюю, среднюю, наружную и заднюю. В ядрах гипоталамуса расположены высшие центры вегетативной нервной системы, причём в ядрах передней группы расположены главным образом центры парасимпатического отдела, а в ядрах задней группы – преимущественно центры симпатического отдела.
В гипоталамусе насчитывается более 30 пар ядер, которые на основе цитоархитектонического строения подразделяют на три не резко разграниченные группы: переднюю, среднюю и заднюю. С функциональной точки зрения ядра гипоталамуса делят на 5 групп: преоптическую, переднюю, среднюю, наружную и заднюю. В ядрах гипоталамуса расположены высшие центры вегетативной нервной системы, причём в ядрах передней группы расположены главным образом центры парасимпатического отдела, а в ядрах задней группы – преимущественно центры симпатического отдела.
В ядрах средней группы находятся центры, регулирующие обмен веществ и пищедобывательные поведенческие реакции. Также в этих ядрах имеются своеобразные нейроны-датчики, реагирующие на различные изменения внутренней среды организма: температуру и вязкость крови, концентрацию минеральных веществ в крови, содержание в ней гормонов и др.
Гипоталамус является интегративным центром регуляции температуры тела. Перерезка мозга у кошки ниже гипоталамуса нарушает терморегуляцию и превращает теплокровное животное в холоднокровное. Повреждение задней группы ядер гипоталамуса вызывает подавление образования тепла в организме и снижение температуры тела - гипотермию. Разрушение передней группы ядер сопровождается нарушением отдачи тепла и повышением температуры организма – гипертермией.
Одной из главнейших функций гипоталамуса является регуляция деятельности гипофиза. Различают две системы регуляции: 1) гипоталамо-аденогипофизарная, состоящая из некоторых ядер средней группы гипоталамуса, которые функционально связаны с аденогипофизом; 2) гипоталамо-нейрогипофизарная, состоящая из некоторых ядер передней группы гипоталамуса, связанных с задней долей гипофиза, т.е. нейрогипофизом.
 Обнаружено, что секреция гормонов аденогипофиза регулируется нейрогормонами гипоталамуса, которые являются как бы гормонами гормонов. Нейрогормоны вырабатываются нейросекреторными клетками вентромедиального и дорсомедиального ядер, входящих в среднюю группу ядер гипоталамуса (рис. 43). Эти ядра образуют так называемую гипофизотропную зону, с которой начинается гипоталамо-аденогипо-физарная система. Нейрогормоны секретируются двух видов: 1) либерины, или рилизинг-факторы, усиливающие секрецию гормонов аденогипофизом; 2) статины (ингибиторы), оказывающие тормозящее действие на выделение некоторых гормонов аденогипофизом. Образующиеся в нейросекреторных клетках нейрогормоны поступают по аксонам этих клеток в кровь и с током крови по кровеносным сосудам поступают из гипоталамуса в аденогипофиз, где воздействуют на клетки, секретирующие тот или иной гормон. Секреция самих либеринов и статинов регулируется по принципу отрицательной обратной связи.
Обнаружено, что секреция гормонов аденогипофиза регулируется нейрогормонами гипоталамуса, которые являются как бы гормонами гормонов. Нейрогормоны вырабатываются нейросекреторными клетками вентромедиального и дорсомедиального ядер, входящих в среднюю группу ядер гипоталамуса (рис. 43). Эти ядра образуют так называемую гипофизотропную зону, с которой начинается гипоталамо-аденогипо-физарная система. Нейрогормоны секретируются двух видов: 1) либерины, или рилизинг-факторы, усиливающие секрецию гормонов аденогипофизом; 2) статины (ингибиторы), оказывающие тормозящее действие на выделение некоторых гормонов аденогипофизом. Образующиеся в нейросекреторных клетках нейрогормоны поступают по аксонам этих клеток в кровь и с током крови по кровеносным сосудам поступают из гипоталамуса в аденогипофиз, где воздействуют на клетки, секретирующие тот или иной гормон. Секреция самих либеринов и статинов регулируется по принципу отрицательной обратной связи.
Гипоталамо-нейрогипофизарная система начинается от нейросекреторных клеток супраоптического и паравентрикулярного ядер передней группы ядер гипоталамуса. В этих клетках образуются гормоны окситоцин и вазопрессин (антидиуретический гормон), которые транспортируются (аксонный транспорт) по их длинным аксонам в нейрогипофиз, где и поступают в кровь. Регуляция секреции окситоцина и вазопрессина осуществляется по механизму нейрогуморального рефлекса, афферентным звеном которого являются нервные пути от осморецепторов или механорецепторов до гипоталамуса, а эфферентным звеном – выделяющийся в кровяное русло гормон.
В гипоталамусе и гипофизе обнаружены энкефалины и эндорфины – особая группа нейрогормонов, которые оказывают на нервные клетки морфиноподобное действие и, по-видимому, играют существенную роль в регуляции вегетативных процессов и поведения.
Благодаря связям гипоталамуса с гипофизом создаётся единая нейрогуморальная регуляция функций.
Участие гипоталамуса в регуляции поведения
При электрическом раздражении разных маленьких участков гипоталамуса у животных наблюдаются разнообразные поведенческие реакции. Важнейшими из таких реакций являются оборонительное поведение и бегство, пищевое поведение (потребление пищи и воды), половое поведение, регуляция сна и бодрствования. Все эти поведенческие комплексы направлены на обеспечение выживания особи и вида и включают в себя моторные, вегетативные и гормональные компоненты.
В гипоталамусе обнаружено несколько зон, обусловливающих пищевое поведение. Так, разрушение вентромедиальных ядер гипоталамуса вызывает чрезмерное потребление пищи и ожирение (рис. 44, Б). В этом случае наблюдаются все реакции, характерные для животного при поиске пищи. Оно начинает есть, даже если не голодно, и при этом пережёвывает несъедобные предметы. Происходит обильное слюноотделение, повышенная перистальтика и усиленное кровоснабжение кишечника. Предполагают, что в этом участке гипоталамуса находится «центр насыщения», регулирующий величину потребления пищи.
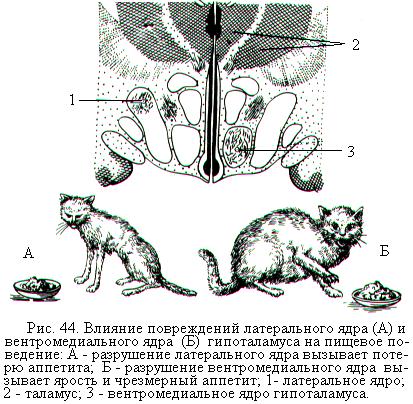 При разрушении в гипоталамусе небольшого участка (латерального ядра), расположенного сбоку от вентромедиальных ядер, наблюдается полный отказ от пищи и воды, истощение и гибель животного (рис. 44, А). Эта зона была названа «центром голода», или «пищевым центром». Если же «центр голода» не разрушать, а возбуждать электрическим раздражением через вживленные электроды, то у животного возникает пищевое поведение, т.е. поиск и приём пищи. Предполагается, что нейроны центра насыщения по мере потребления пищи активируются и подавляют функции центра голода.
При разрушении в гипоталамусе небольшого участка (латерального ядра), расположенного сбоку от вентромедиальных ядер, наблюдается полный отказ от пищи и воды, истощение и гибель животного (рис. 44, А). Эта зона была названа «центром голода», или «пищевым центром». Если же «центр голода» не разрушать, а возбуждать электрическим раздражением через вживленные электроды, то у животного возникает пищевое поведение, т.е. поиск и приём пищи. Предполагается, что нейроны центра насыщения по мере потребления пищи активируются и подавляют функции центра голода.
Гипоталамус также участвует в регуляции приспособительных поведенческих реакций, направленных на поддержание водного баланса организма. В передней области гипоталамуса имеются нейроны с осморецепторной функцией, т.е. обладающие способностью возбуждаться при увеличении осмотического давления крови, вызванного уменьшением воды в организме. Дефицит воды создаёт мощную мотивацию, направленную на его устранение, и появление чувства жажды.
В заднем отделе гипоталамуса обнаружены зоны, связанные с регуляцией полового поведения. В опытах на крысах с хронически вживленными в эти зоны электродами было установлено, что подопытные крысы довольно быстро обучались нажимать на рычаг (рис. 45), который включает электрическое раздражение их собственного гипоталамуса, вызывающее чувство полового удовольствия. Если животным предоставлялся свободный выбор, то голодные крысы гораздо чаще устремлялись к рычагу самостимуляции, чем к лежащей рядом пище, и нажимали на рычаг до 5000 за 1 час.
Работы, проведённые в условиях нейрохирургической клиники, показали, что раздражение аналогичных участков мозга у людей вызывало чувства радости, удовлетворения, сопровождавшиеся эротическими переживаниями. Материалы клинических наблюдений свидетельствуют о том, что гипоталамус регулирует правильную периодичность функций, связанных с размножением. Опухоли в области гипоталамуса могут вызывать быстрое половое созревание, нарушение менструального цикла, половую слабость и ряд других дисфункций.
При локальном электрическом раздражении передних отделов гипоталамуса у бодрствующих животных возникает оборонительное поведение. Например, при таком раздражении кошка принимает угрожающую позу, оскаливает зубы, шипит и выпускает когти. У неё изменяются вегетативные реакции – учащается дыхание, подымается шерсть на спине, расширяются зрачки, возрастает артериальное давление, кровоток в скелетных мышцах увеличивается, а в кишечнике уменьшается. Из мозгового вещества надпочечников в кровь выбрасываются адреналин и норадреналин, а из гипофиза – адренокортикотропный гормон, приводящий к увеличению выделения гормонов корковым веществом надпочечников. В связи с тем, что данная агрессивная реакция не направлена на какой-то объект, то она называется ложной яростью.
При раздражении некоторых боковых отделов гипоталамуса агрессивное поведение животного имеет чёткую направленность против конкретного объекта и осуществляется без лишних движений и демонстрации устрашения.
Области гипоталамуса, раздражение которых приводит к поведенческим реакциям, широко перекрываются. В связи с этим пока не удалось точно определить конкретные группы нейронов или другие анатомические структуры, отвечающие за то или иное поведение.
Гипоталамус принимает участие в чередовании состояний сна и бодрствования. При электрическом раздражении некоторых зон среднего гипоталамуса бодрствующие животные засыпают, а при раздражении заднего гипоталамуса спящие животные просыпаются и поддерживаются в состоянии бодрствования. Известны случаи перехода людей в состояние летаргического сна при повреждении гипоталамуса.
Таким образом, гипоталамус является важным центром регуляции поведенческих реакций. Вместе с тем, нейронная организация гипоталамуса, благодаря которой это маленькое, весом 5 г у взрослого человека, образование способно управлять множеством жизненно важных поведенческих и нейрогуморальных процессов, остаётся мало изученной.
СТРОЕНИЕ И ФУНКЦИИ КОНЕЧНОГО МОЗГА
Большие полушария головного мозга. Подкорковые ядра
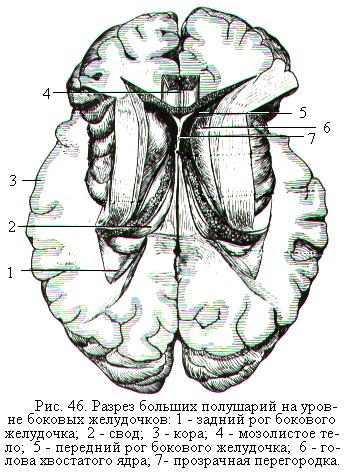
Конечный мозг – это самый крупный и главный отдел головного мозга у человека, управляющий деятельностью других отделов головного мозга и спинным мозгом. Он составляет 80 % массы всего головного мозга человека и состоит из сильно развитых парных частей – правого и левого большого полушария. Большие полушария разделены продольной щелью, в глубине которой соединяются между собой мозолистым телом (рис. 46). Мозолистое тело представляет собой белое вещество, состоящее из нервных волокон, соединяющих правое и левое полушария. В толще белого вещества ниже уровня мозолистого тела в обоих полушариях находятся полости - боковые желудочки (в правом полушарии – правый, или 1-й боковой желудочек, а в левом полушарии – левый, или 2-й боковой желудочек). Они являются местом образования и ёмкостью для цереброспинальной жидкости и сообщаются с 3-м желудочком, расположенным в промежуточном мозге.
Под мозолистым телом находится свод. Он образован двумя изогнутыми пучками нервных волокон, которые в средней части соединены между собой, а спереди и сзади расходятся, образуя столбы и ножки свода. Между передней частью мозолистого тела и сводом натянута тонкая вертикальная пластинка мозговой ткани – прозрачная перегородка (рис. 46). Снаружи большие полушария покрыты тонким слоем серого вещества. Его называют плащом, или чаще корой больших полушарий.
Под корой расположено белое вещество больших полушарий, в глубине которого находятся скопления серого вещества – подкорковые ядра,
 или подкорковые узлы, или базальные ганглии. К подкорковым ядрам относятся хвостатое ядро и отделённое от него полоской белого вещества чечевицеобразное ядро. Чечевицеобразное ядро в свою очередь делится полоской белого вещества на скорлупу и бледный шар, или бледное ядро (паллидум). В состав базальных ганглиев часто включают также ограду и реже – миндалевидное тело, или миндалину. Хвостатое ядро и скорлупа примыкают друг к другу и их часто объединяют под общим названием «полосатое тело» (стриатум) из-за того, что скопления тел нейронов, образующих серое вещество этих ядер, чередуются с прослойками белого вещества.
или подкорковые узлы, или базальные ганглии. К подкорковым ядрам относятся хвостатое ядро и отделённое от него полоской белого вещества чечевицеобразное ядро. Чечевицеобразное ядро в свою очередь делится полоской белого вещества на скорлупу и бледный шар, или бледное ядро (паллидум). В состав базальных ганглиев часто включают также ограду и реже – миндалевидное тело, или миндалину. Хвостатое ядро и скорлупа примыкают друг к другу и их часто объединяют под общим названием «полосатое тело» (стриатум) из-за того, что скопления тел нейронов, образующих серое вещество этих ядер, чередуются с прослойками белого вещества.
|
|
|
|
|
Дата добавления: 2014-12-26; Просмотров: 752; Нарушение авторских прав?; Мы поможем в написании вашей работы!