КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Розпад гема
|
|
|
|
1. Розрив тетрапірольного кільця гему (у складі гемоглобіну) шляхом окислювального розщеплення метинового містка між І та II кільцями протопорфіринового циклу; в результаті реакції червоний пігмент еритроцитів гемоглобін перетворюється на зелений кров'яний пігмент вердоглобін (холеглобін). Реакція каталізується ферментом НАДФН-залежною гемоксигеназою і
супроводжується виділенням монооксиду вуглецю. Перетворення гемоглобіну на вердоглобін внаслідок окислення гему спричиняє послідовну зміну забарвлення в ділянках гематом, що утворюють "синці"
2. Розпад вердоглобіну з відщепленням білкової частини, вивільненням іона заліза та утворенням тетрапірольної молекули білівердину.
3. Перетворення білівердину на білірубін шляхом відновлення метинового зв'язку між піроламиIII, IV. Реакція каталізується ферментом НАДФН-залежною білівер- динредуктазою
4. Зазначені етапи утворення жовчних пігментів (1-3) відбуваються в клітинах ретикулоендотеліальної системи, з яких білірубін надходить у кров, де адсорбується молекулами сироваткового альбуміну. Комплекс "сироватковий альбумін - білірубін" транспортується в печінку, де пігмент поглинається гепатоцитами і підлягає подальшим перетворенням.
5. Білірубін є ліпідорозчинною речовиною і у високих концентраціях проявляє мембранотоксичність, особливо щодо клітин головного мозку. Детоксикація білірубіну, яка полягає в перетворенні пігменту у водорозчинну (і менш токсичну) форму - глю- куронід білірубіну, відбувається в мембранах ендоплазматичного ретикулуму гепатоцитів. У процесі взаємодії білірубіну з УДФ-глюкуроновою кислотою (УДФГК) утворюються моно- та диглюкуроніди білірубіну: Реакція каталізується УДФ-глюкуронілтрансферазою. Основна частина білірубіну екскретується в жовч у формі диглюкуронідів;
4. Біохімія нейромедіаторів та фізіологічно активних сполук.
Нейромедіатори (нейротрансмітери) - біомолекули, які забезпечують передавання імпульсів (хімічних сигналів) у нервовій системі з одного нейрона на інший, а також з нейрона на ефекторний орган. За хімічною природою нейромедіатори поділяють на такі сполуки: ацетилхолін,
біогенні аміни (катехоламіни — норадреналін, дофамін, серотонін), амінокислоти та їх похідні (у-аміномасляна кислота - ГАМК, гліцин, глутамат, аспартат), пептиди - нейропептиди (ендорфіни, енкефаліни, сполука Р тощо). Медіаторні функції в нервовій системі можуть також виконувати пролін, таурин, (З-аланін, аденозин, простагландини. Рецептори нейромедіаторів - мембранні білки (здебільшого глікопротеїни), що локалізовані в постсинаптичних мембранах нейронів або плазматичних мембранах клітин ефекторних органів і здатні до зв'язування фізіологічних ефекторів (нейромедіаторів, різних ФАС, психотропних сполук) і передавання зовнішньоклітинного хімічного сигналу всередину нейрона. За принципами молекулярної організації та функціонування, рецептори нейромедіаторів - це здебільшого іонотропні рецептори (рецептори І класу), тобто такі, що контролюють відкриття іонних каналів на мембрані для Са2+, Na+ і К+; в ролі первинних ефекторів, що передають хімічний сигнал на нервову клітину, в цьому разі виступають компоненти іонних каналів (розділ 23). Крім того, у фізіологічних ефектах
деяких нейромедіаторів та нейромодуляторів (зокрема, нейропептидів головного мозку, ацетилхоліну та деяких біогенних амінів) беруть участь і метаботропні рецептори (рецептори II класу), які активують внутрішньоклітинні біохімічні системи шляхом утворення цАМФ або цГМФ, включенням фосфоінозитидної системи та/або збільшенням цитозольної концентрації іонів Са2+. Крім нейромедіаторів, із зазначеними типами рецепторів можуть взаємодіяти численні лікарські засоби, нейротоксини, що активують (агоністи), гальмують, блокують (антагоністи) або модулюють біохімічні, нейрофізіологічні та психологічні (поведінкові) ефекти, опосередковані
збудженням специфічних рецепторів певних зон головного мозку.
5. Роль печінки в обміні білків, жирів та вуглеводів
/. Вуглеводна (глікогенна) функція печінки Ця функція полягає в здатності гепатоцитів утворювати лабільні резерви вуглеводів, що використовуються для підтримання необхідних концентрацій глюкози в крові та постачання цього цукру в інші органи (насамперед, головний мозок) у періоди між прийомами їжі. Утворення й утилізація глюкозо-6-фосфату
Крім використання ліпідів для власних енергетичних та структурних потреб, печінці належить визначальна роль у регуляції окислення жирів іншими тканинами - функція, що реалізується шляхом утворення в гепатоцитах і секреції в кров триацилглщеролів (у формі лїпопротеїнів дуже низької щільності — ЛПДНЩ) та кетонових тіл. У печінці утворюється також основна частина холестерину, що використовується в периферичних тканинах для синтезу фізіологічно активних стероїдів. Роль печінки в білковому обміні цілісного організму полягає в утворенні більшості
білків плазми крові, які виконують важливі біохімічні та фізіологічні функції, регуляції розподілу амінокислот між окремими органами і тканинами та синтезі сечовини як кінцевого продукту азотистого катаболізму.
6. Потреба в ліпідах, роль лейкотрієпів, простагландинів, тромбоксанів. Ліпіди і захворювання.
Ліпіди - клас біоорганічних сполук, характерною ознакою яких є нерозчинність у воді й інших полярних розчинниках та здатність до розчинення в неполярних (гідрофобних) рідинах. Неполярні розчинники (діетиловий ефір, тетрахлорметан, хлороформ тощо) використовують для екстрагування ліпідів із біологічних об'єктів (крові, тканин т.і.). В організмі людини та тварин ліпіди виконують важливі функції як акумулятори та постачальники енергії, компоненти структури клітин, особливо біологічних мембран; певні представники ліпідів є фізіологічно активними речовинами (вітаміни, гормони). Лейкотрієни - гідроксипохідні арахідонової кислоти, спряжені трієни, що, на відміну від інших ейкозаноїдів, не містять у собі циклічної структури. Залежно від особливостей хімічної будови, розрізняють декілька типів лейкотрієнів. Простагландини та простацикліни — гідрокси- похідні 20-вуглецевих жирних кислот, що містять у своїй структурі п'ятичленний цикл. Разом ці сполуки складають підклас простаноїдів — похідних простанової кислоти, яка утворюється за рахунок замикання зв'язку між 8-м та 12-м
вуглецевими атомами в молекулі арахідонової (С204) кислоти. Простагландини позначаються скорочено ПГ (PG - Prostaglandin, англ.) з додаванням заголовної літери латинського алфавіту (А, В, D, Е, F, G, Н, І), цифрового індексу, що вказує на кількість подвійних звязків у бічному
ланцюзі, та (в деяких випадках) літери грецького алфавіту (позначає певний ізомер). Довжина бічних ланцюгів R у більшості простагландинів складає 7 або 8 вуглецевих атомів. Окремі представники простагландинів розрізняються наявністю та розташуванням кето- або гідроксильної групи в кільці або бічному ланцюгу, будовою бічних ланцюгів (ЬЦ, R8, R4), наявністю в них подвійних зв'язків. Своєрідну будову з наявністю внутрішньої циклічної кисневої структури має простагландин PGI2, що отримав назву простацикліну (рис. 25.9).
2. Тромбоксани - гідроксипохідні 20-вуглецевих жирних кислот, що містять у своїй структурі 6-членний кисеньвмісний цикл. Активна форма тромбоксанів - тромбоксани А - мають внутрішній атом кисню в гетероциклічному кільці.
Тромбоксани є фізіологічними антагоністами антитромботичних ефектів прос- тацикліну. На відміну від простацикліну, тромбоксани, особливо тромбоксан А2, що також утворюється в інтимі кровоносних судин, спричиняють скорочення гладеньких м'язів судин та сприяють агрегації тромбоцитів. Біохімічні механізми проагрегантної дії тромбоксану полягають у його позитивному впливі на мобілізацію з внутрішньоклітинних депо іонів Са2+, які спричиняють активацію скорочувальних білків тромбоцитів та їх адгезію до поверхні ендотелію.
4. Важливою функцією ейкозаноїдів різних класів, особливо простагландинів, лейкотрієнів та ліпоксинів, є їх участь у розвитку і регуляції такого загально- патологічного процесу, як запалення, яке є біологічним захистом тканин від дії пошкоджуючих факторів.
Стимулюють розвиток запального процесу – простагландини.
Білет 8
1. Етапи та механізми трансляції. Ініціація. Елонгація. Термі нація.
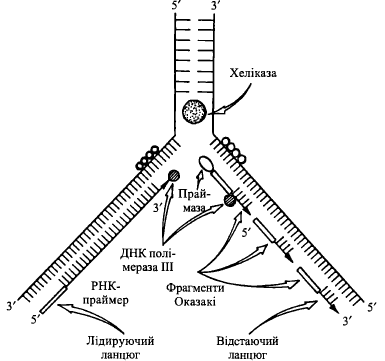 1. Ініціація синтезу полідезоксирибонуклеотидних ланцюгів ДНК, якій передує утворення "затравних" (праймерних) ланцюгів РНК (РНК-праймерів), до 3'- ОН-груп яких здатні приєднуватися дНМФ, що утворюють нові (дочірні) ланцюги ДНК. Довжина цих праймерних ланцюгів складає в середньому від 10 до 200 нуклеотидів. Синтез РНК-праймерів відбувається за участю ферментів РНК-полімераз - так званих праймаз.
1. Ініціація синтезу полідезоксирибонуклеотидних ланцюгів ДНК, якій передує утворення "затравних" (праймерних) ланцюгів РНК (РНК-праймерів), до 3'- ОН-груп яких здатні приєднуватися дНМФ, що утворюють нові (дочірні) ланцюги ДНК. Довжина цих праймерних ланцюгів складає в середньому від 10 до 200 нуклеотидів. Синтез РНК-праймерів відбувається за участю ферментів РНК-полімераз - так званих праймаз.
2. Елонгація синтезу ДНК, яка відбувається за різними механізмами на лідируючому та відстаючому ланцюгах.
2.1. На лідируючому ланцюгу нарощування дНМФ здійснюється ДНК-поліме- разою III, що функціонує безперервно, утворюючи ланцюг ДНК від РНК-праймера до реплікативної вилки.
2.2. На відстаючому ланцюгу: а) спочатку при дії ДНК-полімерази III синтезуються окремі фрагменти Оказакі, кожен з яких починається з відповідного РНК-праймера і закінчується перед
початком передуючого йому РНК-праймера; б) після формування фрагментів Оказакі ДНК-полімераза І, за рахунок своєї 5'—>3'- екзонуклеазної активності, видаляє РНК-праймери і (за рахунок 5'—»3'-полімеразної активності) заміщує їх фрагментами ДНК;
в) розриви між окремими фрагментами Оказакі зшиваються спеціальним ферментом ДНК-лігазою.
2. Гормони гіпофіза
Передня частка гіпофіза (аденогіпофіз) продукує значну кількість гормонів, які стимулюють фізіологічні та біохімічні процеси в різних тканинах-мішенях, у тому числі активують дію інших ендокринних залоз (тропна функція гіпофізарних гормонів). Виходячи з особливостей молекулярної генетики, біосинтезу та структурно-функціональних властивостей, гормони аденогіпофіза утворюють три групи: I група - "гормон росту-пролактин-хоріонічний соматомамотропін"; II група - глікопротеїни - "тропні гормони гіпофіза"; III група - похідні "проопіомеланокортину"
1. Гормон росту (соматотропін, соматотропний гормон, СТГ) - простий білок, що складається з одного поліпептидного ланцюга. Головна функція СТГ - стимуляція постнатального росту організму; ця складна біологічна функція реалізується за рахунок різноманітного спектра впливу гормону на біосинтез білка, вуглеводний та ліпідний метаболізм. 2. Пролактин (лактогенний гормон, мамотропін, лютеотропний гормон) - простий білок, що складається з одного поліпептидного ланцюга. 3. Хоріонічний соматомамотропін {ХС; плацентарний лактоген) -гормон, що проявляє лактогенну та лютеотропну активності, а за метаболічними ефектами близький до соматотропіну.
2. включає в себе сполуки глікопротеїнової природи: тиреотропний та гонадотропний гормони гіпофіза і плаценти. Тиреотропний гормон (ТТГ; тиротропін) - подібно до інших глікопротеїнових гормонів гіпофіза є димером типу альфа-бета. Основна біологічна функція ТТГ - підтримка функціональної активності (синтезу тиреоїдних гормонів) та структури щитовидної залози. Гонадотропні гормони: 1. Фолікулостимулюючий гормон {ФСГ; фолітропін) — білок з м.м. 33 кДа. Мішенями для ФСГ є фолікулярні клітини яєчників та клітини Сертолі сім'яників. Передача хімічного сигналу при дії ФСГ здійснюється за рахунок активації аденілатциклази. 2. Лютеїнізуючий гормон (ЛГ; лютропін; гормон, що стимулює інтерстиці- альні клітини Лейдига - ГСІК) - білок з м.м. 29 кДа. Рецептори для ЛГ локалізовані на плазматичних мембранах клітин яєчників (у жіночому організмі) та клітин Лейдига сім'яників (у чоловічому організмі). Як і в разі ФСГ, вторинним месендже- ром у дії ЛГ на ефекторні системи клітин є цАМФ. 3. Хоріонічний гонадотропін (ХГ) - білок з м.м. близько 37 кДа; синтезується трофобластом плаценти. Біологічна роль гонадотропінів полягає в регуляції функцій статевої сфери людини як в препубертатному та пубертатному періодах, так і у дорослих особин, процесів як гаметогенезу, так і продукції статевих гормонів.
3. Адренокортикотропний гормон (АКТГ; кортикотропін) - одноланцюговий пептид, що складається з 39 амінокислотних залишків (м.м. 4,5 кДа). Головними мішенями АКТГ є клітини кори надниркових залоз, відносно яких гормон проявляє два типи біологічної активності: стимуляцію стероїдогенезу та підтримання маси надниркових залоз. 2. Ліпотропний гормон (ЛПГ; ліпотропін) - група пептидів, що мають властивості активувати ліполіз в адипоцитах жирової тканини і мобілізацію жирних кислот. 3. Ендорфіни - представники групи опіоїдних нейропептидів (а-, (3-, у- та д-ендорфіни), що виконують функції нейромедіаторів, ендогенних знеболювальних факторів та модуляторів певних важливих психофізіологічних процесів у пептидергіч- них структурах головного мозку (розділ 33). 4. Меланоцитостимулюючий гормон (МСГ) — група пептидів (а-, Р-, у-МСГ), які продукуються у проміжній частці гіпофіза і стимулюють функціональну активність меланоцитів шкіри, збільшуючи її пігментацію.
Гормони задньої частки гіпофіза – вазопресин і окситоцин. Накопичуються, у нейрогіпофізі. Впливають на нирки.
|
|
|
|
|
Дата добавления: 2015-05-08; Просмотров: 1461; Нарушение авторских прав?; Мы поможем в написании вашей работы!