КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Для переноса белков из цитозоля во внешнюю митохондриальную мембрану также необходимо их
|
|
|
|
8-17
8.4.5. Для транспорта белков в межмембранное пространство митохондрий необходимы два сигнала [25]
Транспорт некоторых белков-предшественников в межмембранное пространство митохондрий начинается с их переноса в матрикс (рис.
8-29). Однако за N-концевым сигнальным пептидом, инициирующим этот перенос, расположена очень сильно гидрофобная аминокислотная последовательность. Как только сигнальный пептид отщепляется матриксной протеазой, эта гидрофобная последовательность начинает, в свою очередь, выполнять роль сигнального пептида для обратного встраивания данного белка во внутреннюю мембрану. Вероятно, этот перенос происходит с помощью механизма, сходного с механизмом встраивания белков в мембрану ЭР. Аналогичный способ используется и для встраивания во внутреннюю мембрану митохондрий белков, кодируемых митохондриальным геномом (рис. 8-30, А).
После того, как белки, предназначенные для межмембранного пространства, встроятся во внутреннюю мембрану, они отрезаются протеазой в межмембранном пространстве (рис. 8-30, Б}. Многие из этих зрелых растворимых белков в конце концов присоединяются к «внешней»
поверхности внутренней мембраны, где они образуют субъединицы комплексов, содержащих также и трансмембранные белки.
Для транспорта белков из цитозоля во внутреннюю мембрану митохондрий также требуется гидрофобный сигнальный пептид.
Возможно, этот транспорт происходит по схеме, изображенной на рис. 8-30, А, но прямо это доказано не было. Такой двухстадийный путь трудно отличить в эксперименте от альтернативного пути, при котором транспорт в точке контакта прерывается по достижении гидрофобного сигнала, и белок остается заключенным в бислой внутренней мембраны.
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
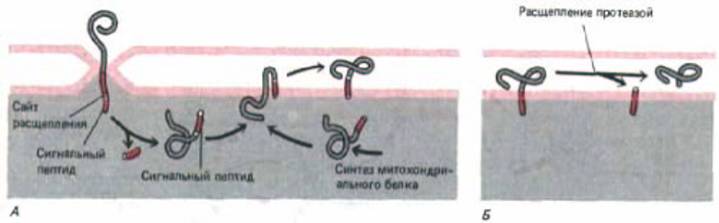
Рис. 8-30. Импорт белков из цитозоля в межмембранное пространство митохондрий или внутреннюю мембрану требует многих сигналов. Вначале белок переносится в пространство матрикса, как это показано на рис. 8-29. Однако при отрезании сигнального пептида, использованного для первичного переноса, обнажается смежный гидрофобный сигнальный пептид на новом N-конце. Этот сигнал вызывает встраивание белка во внутреннюю мембрану таким же образом, каким в нее встраиваются белки, кодируемые митохондриальным геномом (А.) Этот механизм предположительно сходен с тем, который бактериальные предки митохондрий использовали для встраивания белков в плазматическую мембрану, и, как полагают, напоминают механизм встраивания белков в ЭР. Для транспорта белков в межмембранное пространство требуется еще третья стадия, на которой протеаза с активным центром, обращенным в трансмембранное пространство, отрезает белок от его трансмембранного сигнального пептида, находящегося во внутренней мембране (Б). Путь, изображенный на рис. (А), может быть использован и для переноса белков из цитозоля во внутреннюю мембрану, который тоже требует гидрофобного пептида.
разворачивание [26]
Во внешней мембране митохондрий имеется одна необычная структура (она напоминает внешнюю мембрану грамотрицательных бактерий), липидный слой которой содержит большие количества образующего поры белка-порина. По этой причине внешняя мембрана свободно проницаема для неорганических ионов и метаболитов и для молекул белков размером меньше 10 кДа. Но для больших по размеру белков внешняя мембрана является барьером и поэтому помогает удержать белки межмембранного пространства от утечки обратно в цитозоль.
Включение во внешнюю мембрану белков, кодируемых основным клеточным геномом - таких, как порин, - происходит по АТР-
зависимому механизму, но не требует использования специальных пептидов, которые бы потом отрезались. Мембранный потенциал тоже не требуется. О том, как происходит подобное включение, известно очень мало. По крайней мере у одного белка внешней мембраны имеется нормальный матриксный транспортный сигнал, за которым следует последовательность, каким-то образом прерывающая перенос на внешней мембране.
|
|
|
|
|
Дата добавления: 2015-08-31; Просмотров: 683; Нарушение авторских прав?; Мы поможем в написании вашей работы!