КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Перенос веществ в митохондриальный матрикс зависит как от электрохимического градиента на внутренней
|
|
|
|
мембране, так и от гидролиза АТР [22]
Практически все сведения о молекулярном механизме переноса белков внутрь митохондрии были получены при анализе бесклеточных транспортных систем. Суть экспериментов заключается в следующем. Вначале из гомогенизированных клеток методом дифференциального центрифугирования выделяют митохондрии, а затем инкубируют их с радиоактивно меченными белками, предназначенными для этих органелл (митохондриальные белки-предшественники). Очищенные белки-предшественники очень быстро и эффективно включаются в такие митохондрии.
Все формы направленного движения и транспорта нуждаются в энергии. В большинстве случаев эта энергия используется в форме АТР.
Однако для переноса белков в митохондрии требуется еще наличие электрохимического градиента на внутренней митохондриальной мембране.
Этот градиент образуется в процессе транспорта электронов по мере того, как протоны откачиваются из матрикса в межмембранное пространство (см. разд. 7.1.7). Внешняя митохондриальная мембрана свободно проницаема для ионов, поэтому на ней не поддерживается никакой градиент.
Электрохимический градиент на внутренней мембране используется как аккумулятор энергии для осуществления большей части синтеза АТР в клетке. Кроме того, энергия градиента расходуется для переноса внутрь митохондрии белков, несущих положительно заряженные митохондриальные сигнальные пептиды. Если добавить ионо-форы, сбрасывающие митохондриальный мембранный потенциал (см. разд. 7.2.10), этот перенос блокируется. Каким образом электрохимический градиент способствует переносу белков? Ответ на этот вопрос пока не получен.
8.4.3. Митохондриальные белки проникают в матрикс в зонах слипания, связывающих две мембраны [23]
Пересекает ли белок на пути к митохондриальному матриксу две мембраны поочередно, или он проникает через обе мембраны сразу?
Чтобы ответить на этот вопрос, можно охладить бесклеточную систему до температуры льда, задержав таким образом белки на промежуточ-
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
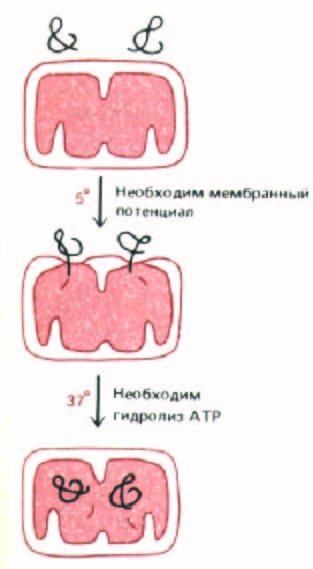
Рис. 8-28. При переносе белков в матрикс они в течение короткого времени соединяют внутреннюю и внешнюю митохондриальные мембраны.
Если изолированные митохондрии инкубировать с белком-предшественником при 5°С, предшественник переносится лишь частично. В матриксе N-
концевой сигнальный пептид отрезается; большая часть полипептидной цепи остается вне митохондрии (и доступна для протеолитических ферментов). При нагревании до 37°С перенос происходит полностью. Для изначального внедрения белка в митохондриальную мембрану при 5°С требуется разность потенциалов на внутренней мембране. Последующий перенос может происходить без этой разности потенциалов, но для него необходимо присутствие АТР с цитоплазматической стороны внутренней мембраны. Полагают, что гидролиз АТР необходим при разворачивании полипептидной цепи для того, чтобы белок мог пройти сквозь мембрану.
ной стадии переноса. Оказалось, что их N-концы находятся при этом в матриксе (они могут быть удалены протеазой матрикса), а остальная часть молекул расположена вне митохондрии (поскольку чувствительна к добавленным извне протеолитическим ферментам). Этот результат показывает, что белок-предшественник, когда проникает в матрикс, проходит через обе митохондриальные мембраны сразу. Специалисты по электронной микроскопии заметили многочисленные зоны слипания, в которых, внешняя и внутренняя митохондриальные мембраны сливаются, и предположили, что это именно те участки, через которые происходит перенос белков в матрикс. Недавно эти точки контакта были идентифицированы биохимически (по их связыванию с частично перенесенными внутрь митохондрии белками-предшественниками) и очищены.
Если охлажденные митохондрии, содержащие частично перенесенные промежуточные продукты, опять нагреть, то перенос быстро завершается (рис. 8-28), даже если мембранный потенциал на внутренней мембране сброшен. По-видимому, мембранный потенциал необходим лишь для начальной стадии переноса белка через мембрану, которая происходит даже при низкой температуре. Дальнейшие события, однако, требуют наличия АТР. Эти факты означают, что в норме перенос проходит в два этапа: 1) управляемое электрически проникновение сигнального пептида и связанных с ним последовательностей сквозь обе митохондриальные мембраны и 2) продвижение остатка цепи в митохондриальный матрикс, требующее гидролиза АТР и физиологических температур (рис. 8-29).
8.4.4. Когда белки проникают в митохондриальный матрикс, они разворачиваются [24]
По всей вероятности, белки-предшественники разворачиваются перед тем, как пересечь две митохондриальные мембраны в точке контакта. Трудно представить себе, что свернутый водорастворимый белок мог бы «протаранить» два (или даже один) липидных бислоя, оставаясь в своей нативной трехмерной конформации. Точно также невозможно вообразить, что пора могла бы пропускать глобулярные белки, которые сильно варьируют по размерам и форме. Ведь при этом она становилась бы проницаемой для протонов, и в результате электрохимический градиент на внутренней мембране исчезал бы. Между тем, в развернутом состоянии все белки имеют сходную конформацию и могут быть перенесены с помощью общего механизма. Но поскольку белки в свернутом состоянии обладают меньшей свободной энергией, чем в развернутом (по этой причине полипептиды спонтанно сворачиваются), разворачивание молекулы белка требует затрат энергии. Предполагается, что эту энергию обеспечивает гидролиз АТР.
Чтобы проверить, разворачиваются ли белки-предшественники, когда они пересекают митохондриальную мембрану, был сконструирован гибридный ген, кодирующий «смешанный» белок. В этом как бы состыкованном белке митохондриальный сигнальный пептид присоединен к N-
концу цитоплазматического фермента-дигидрофолатредуктазы (ДГФР). Такой гибридный белок обладал почти ненарушенной ферментативной активностью, а значит ДГФР была в нативной трехмерной конформации. При смешивании с препаратом митохондрий этот белок поступал в матрикс. Однако если его предварительно обрабатывали метатрексатом, который прочно связывается с активным центром фермента и мешает разворачиванию его молекулы, то перенос резко подавлялся.
Генетические эксперименты на дрожжах подтверждают, что для
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
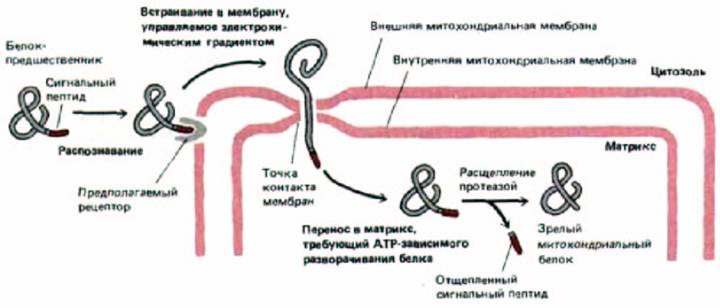
Рис. 8-29. Импорт белков в митохондрии. N-концевой сигнальный пептид белка-предшественника распознается рецептором, который, как полагают, расположен во внешней мембране. Белок переносится через обе митохондриальные мембраны в специальных точках контакта. Для начала этого процесса необходим электрохимический градиент по сторонам внутренней мембраны. В матриксе сигнальный пептид отрезается специфической протеазой, и образуется зрелый белок.
АТР-зависимой реакции разворачивания белков необходимы некоторые гены семейства hsp 70. Если эти гены инактивировать, то и митохондриальные белки-предшественники, и белки, предназначенные для ЭР, не могут пересечь соответствующие мембраны и вместо этого накапливаются в цитозоле.
|
|
|
|
|
Дата добавления: 2015-08-31; Просмотров: 476; Нарушение авторских прав?; Мы поможем в написании вашей работы!