КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
У некоторых трансмембранных белков вскоре после их поступления в ЭР С-концевой трансмембранный
|
|
|
|
участок обменивается на ковалентно связанный фосфолипид инозитол [44]
Как обсуждалось ранее, некоторые ферменты цитозоля катализируют ковалентное присоединение к определенным белкам единичной жирной кислоты (см. разд. 8.2.3). Недавно было обнаружено, что сходный процесс имеет место и в ЭР: карбоксильный конец некоторых белков плазматической мембраны с помощью специфических ферментов ковалентно присоединяется к остатку сахара в гликолипиде. Механизм образования этой связи представлен на рис. 8-55. Установлено, что при этом к белку добавляется гликозилированная молекула фосфатидилинозитола, содержащая две жирных кислоты. Такая модификация обнаружена для большого числа белков плазматической мембраны, включая одну из форм адгезивного белка нейронов и главный белок оболочки трипаносомы. Поскольку оба эти белки связаны с плазматической мембраной только указанным выше способом, в принципе они могут отделяться от клетки в растворимой форме в ответ на сигнал, активирующий специфическую фосфолипазу в плазматической мембране, однако до сих пор подтвердить эту гипотезу экспериментально не удалось.
8.6.14. Большая часть липидных бислоев мембран собирается в ЭР [45]
В мембране ЭР образуются почти все липиды, необходимые для построения новых клеточных мембран, включая фосфолипиды и холестерол. Основной синтезируемый фосфолипид - это фосфатидилхолин (называемый еще лецитином), который может образовываться в три этапа из двух жирных кислот, глицерофосфата и холина (рис. 8-56). Каждый этап катализируется в мембране ЭР ферментами, активные центры которых обращены в цитозоль (именно там находятся все необходимые метаболиты). На первом этапе ацилтрансфераза добавляет к глицерофосфату две жирных кислоты с образованием фосфатидиловой кислоты, соединения достаточно гидрофобного, чтобы остаться после синтеза в липидном бислое. Именно на этом этапе липидный бислой увеличи-
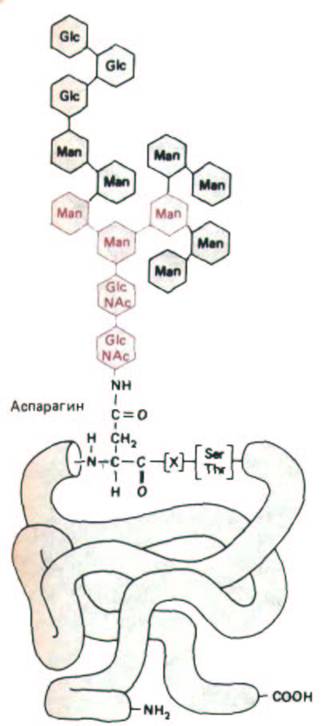
Рис. 8-52. Структура связанного с аспарагином олигосахарида, который добавляется к большинству белков на внутренней стороне мембраны ЭР. Сахара, выделенные цветом, образуют «сердцевину», или кор, этого олигосахарида. Во многих гликопротеинах после разнообразной «доделки» олигосахарида в аппарате Гольджи в нем остаются от первоначальной структуры только сахара, составляющие кор (см. рис. 8-63).
Обратите внимание, что аспарагин находится в последовательности Asp-X-Ser или Asp-X-Thr, где X может быть любой аминокислотой, кроме пролина.
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.

Рис. 8-53. N-связанное гликозилирование белков в ЭР. Почти тотчас после того, как полипептидная цепь попадает в просвет ЭР, она гликозилируется по доступным остаткам аспарагина. Олигосахарид, показанный на рис. 8-52, переносится к аспарагину как целая единица; эту реакцию катализирует связанный с мембраной фермент гликозил-трансфераза.
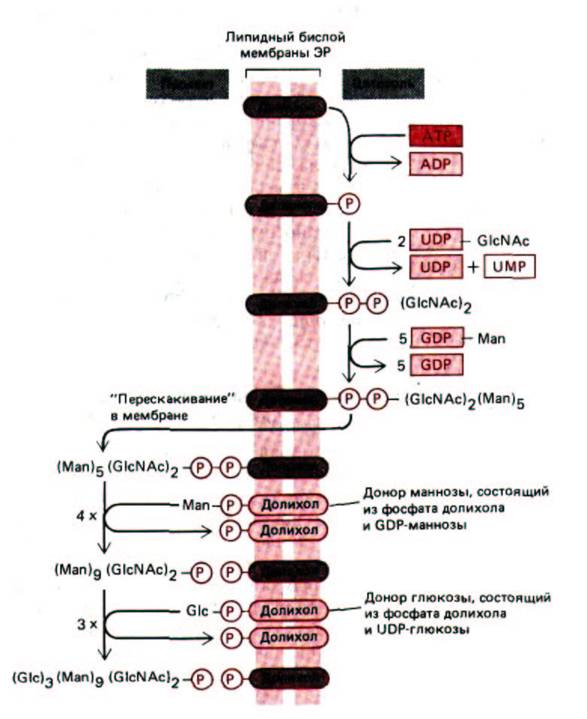
Рис. 8-54. Синтез липид-связанного олигосахарида, который переносится к остаткам аспарагина на внутренней стороне мембраны ЭР.
Этот олигосахарид собирается сахар за сахаром на каркасе из молекулы липида долихола (полиизопреноид-см. схему 2-4). Первый сахар присоединяется к долихолу пирофосфатным мостиком. Эта высокоэнергетическая связь затем активирует олигосахарид для переноса его от молекулы липида к боковой цепи аспарагина. Синтез олигосахарида начинается на цитозольной стороне мембраны ЭР. После того, как промежуточный продукт -липид-MansGlcNAc2 «перепрыгнет» через мембрану, синтез продолжается на внутренней ее стороне. Сокращения: GlcN Ac-N-ацетил глюкозамин, Man -манноза, Glc- глюкоза.
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
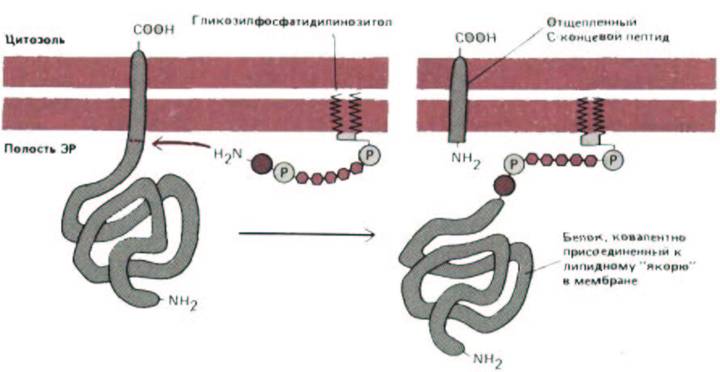
Рис. 8-55. Синтез белков, связанных с мембраной при помощи «якоря» из фосфатидилинозитола. Сразу после завершения синтеза белок остается связанным с мембраной только своим гидрофобным С-концом, состоящим из 15-20 аминокислот, а остальная часть молекулы находится в просвете ЭР. Меньше чем через минуту фермент в ЭР отрезает белок от его мембранной С-концевой части, и одновременно новый карбоксильный конец присоединяется к образовавшемуся ранее интермедиату -гликозилфосфатидилинозитолу. За счет этого ковалентно присоединенного липидного якоря белок остается связанным с мембраной. Все его аминокислоты находятся на внутренней стороне мембраны ЭР, а если белок будет транспортирован к плазматической мембране, они окажутся обращенными во внеклеточное пространство. Точная структура этого гликолипидного головного участка неизвестна.
вается. На последующих стадиях формируется «голова» вновь образованной молекулы липида и, следовательно, химическая природа бислоя, но роста мембраны по объему при этом не происходит (см. рис. 8-56). Все основные фосфолипиды мембраны - фосфатидилхолин (ФХ), фосфатидилэтаноламин (ФЭ), фосфатидилсерин (ФС) и фосфатидилинозитол (ФИ) - синтезируются таким способом.
Как исходное образование фосфатидиловой кислоты, так и ее последующие модификации с формированием различных типов молекул фосфолипидов происходят в той половине липидного бислоя ЭР, которая обращена к цитозолю. Этот процесс мог бы в конце концов превратить липидный бислой в монослой, если бы не существовало механизма для переноса части вновь образованных молекул фосфолипидов в другую половину бислоя ЭР. В искусственных липидных бислоях липиды не совершают таких «флип-флоп»-переходов. В ЭР же количество фосфолипидов выравнивается с двух сторон мембраны за минуты, что почти в 100000 раз быстрее, чем скорость, рассчитанная для спонтанного «флип-флопа».
Полагают, что столь быстрое перемещение поперек бислоя происходит при участии транслокаторов фосфолипидов, которые специфичны для каждого их типа (в зависимости от головной группы). По-видимому, в мембране ЭР имеется транслокатор («флип-паза»), который способен переносить холин-содержащие фосфолипиды (но не этаноламин-, серин- или инозитол-содержащие) из одной половины бислоя в другую. Это означает, что ФХ достигает внутренней поверхности бислоя гораздо легче, чем ФЭ, ФС или ФИ. Таким образом транслокатор отвечает за асимметричное расположение липидов в бислое (рис. 8-57).
Известно, что в ЭР образуются также холестерол и церамид. Церамид экспортируется в аппарат Гольджи, где он служит предшественником двух типов липидов: к одним молекулам церамида присоединяются олигосахаридные цепи с образованием гликосфинголипидов, а к другим - головная фосфохолиновая группа от фосфатидилхолина, и получается сфингомиелин. Таким образом, и гликолипиды, и сфингомиелин в процессе формирования мембран образуются сравнительно поздно. Расположены они исключительно в нецитозольной половине липидного бислоя, поскольку именно там находятся синтезирующие их ферменты.
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.

Рис. 8-56. Синтез фосфолипидов протекает на цитоплазматической стороне мембраны ЭР. Каждый фермент, участвующий в этом синтезе, представляет собой интегральный мембранный белок ЭР, активный центр которого обращен к цитозолю. В цитозоле есть все соединения, необходимые для сборки фосфолипидов. В процессе, изображенном здесь, из комплекса жирная кислота - кофермент А, глицерол-3-фосфата и цитидиндифосфохолина образуется фосфатидилхолин (CDF-холин).
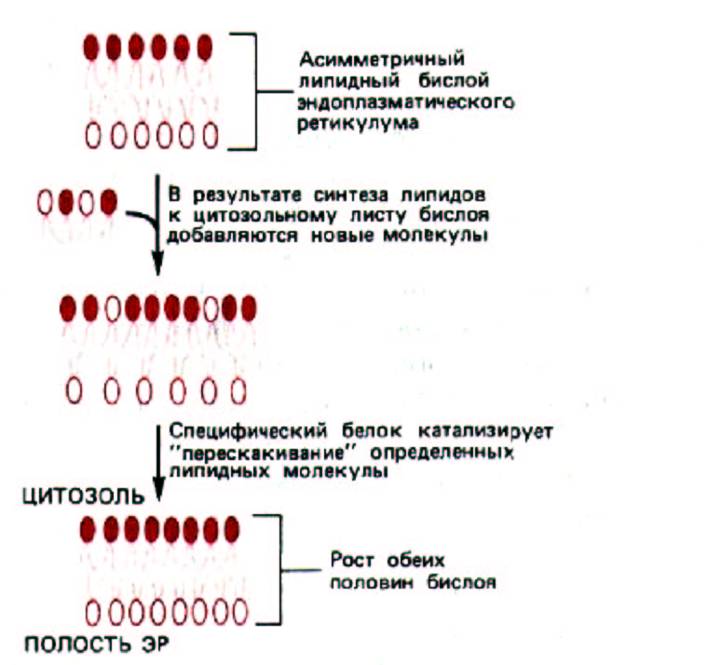
Рис. 8-57. Рост обеих половин липидного бислоя мембраны ЭР требует каталитического «флиппинга» молекул фосфолипидов из одного монослоя в другой. Так как новые молекулы липидов добавляются только к цитоплазматическому монослою и липиды не перескакивают из одного монослоя в другой спонтанно, требуются связанные с мембраной переносчики фосфолипидов («флиппазы»), чтобы переносить определенные молекулы липидов во внутренний слой мембраны. В результате мембрана растет равномерно, как бислой. Поскольку эти ферменты избирательно узнают и переносят только некоторые типы липидов, в ЭР образуется асимметричный бислой. В частности, внутренний слой (из которого образуется внешняя половина бислоя плазматической мембраны) обогащен фосфатидилхолином.
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
|
|
|
|
|
Дата добавления: 2015-08-31; Просмотров: 969; Нарушение авторских прав?; Мы поможем в написании вашей работы!