КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Для ресничек и жгутиков характерны колебательные движения - волны изгиба
|
|
|
|
Реснички движутся координированно, образуя на поверхности клетки однонаправленные бегущие волны (рис. 11-49). Каждая ресничка работает подобно крошечному хлысту: активный удар вперед, при котором ресничка полностью выпрямляется и преодолевает сопротивление окру-
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.

Рис. 11-49, Микрофотографии ресничного эпителия из кишки морского червя, полученные с помощью панирующего электронного микроскопа. Хотя реснички распределены по всей поверхности равномерно, их биение координируется таким образом, что возникают ряды бегущих в одном направлении волн. Эти волны более отчетливо видны при меньшем увеличении справа).
жающей жидкости, сменяется пассивной фазой, во время которой она возвращается в исходное положение, изгибаясь при этом так, чтобы уменьшить сопротивление среды (рис. 11-50, А). Циклы соседних ресничек едва заметно сдвинуты во времени, что приводит к волнообразной картине, которую можно видеть под микроскопом.
Жгутики спермиев и многих простейших по своей внутренней структуре очень похожи на реснички, но обычно они гораздо длиннее, и характер движения у них иной: не удары хлыста, а бегущие квазисинусоидальные волны (рис. 11-50, Б). Тем не менее молекулярная основа движения здесь одна и та же. Следует, однако, отметить, что жгутики бактерий - это нечто совсем иное, чем реснички и жгутики эукариотических клеток (см. разд. 12.5.4).
11.3.2. Ресничка содержит пучок параллельных микротрубочек, образующих структуру типа 9+2
Биение реснички обусловлено изгибанием ее осевой структуры - так называемой аксонемы. Это сложный комплекс микротрубочек и связанных с ними белков. Микротрубочки обычно представляют собой полые белковые цилиндры с наружным диаметром 25 нм (см. ниже). В аксономе они видоизменены и расположены весьма характерным образом. Открытие этой системы явилось одним из самых впечатляющих результатов ранних электронно-микроскопических исследований: девять сдвоенных трубочек расположены по окружности, в центре которой находятся две одиночные микротрубочки (рис. 11-51). Такая структура типа 9 + 2 характерна для ресничек и жгутиков почти всех эукариотических организмов, от простейших до человека. Микротрубочки тянутся
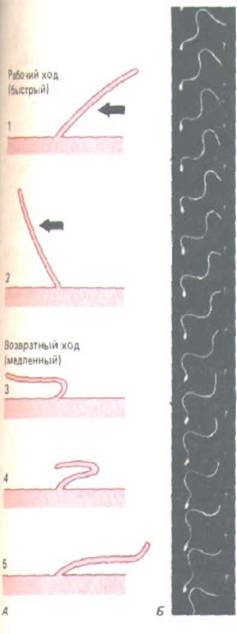
Рис. 11-50. Различный характер биения ресничек и жгутиков. А. Удар реснички (например, реснички эпителия дыхательных путей)
напоминает гребное движение пловца. За рабочим ходом (стадии 1 и 2), при котором жидкость прогоняется по поверхности клетки, следует совершенно иной по характеру возвратный ход (стадии 3, 4, 5). Каждый цикл длится обычно 0,1-0,2 с и создает силу, перпендикулярную оси аксонемы. Б. Здесь для сравнения показано волнообразное движение жгутика у спермия оболочника. Клетка была заснята при стробоскопическом освещении с частотой 400 вспышек в 1 с. Обратите внимание, что волны, имеющие постоянную амплитуду, непрерывно движутся от основания жгутика к его концу. В результате клетка движется прямо вперед, как бы отталкиваясь от своей аксонемы, т.е. совершенно иначе, чем в случае с ресничкой. (Б-с любезного разрешения С. J. Brokaw.)
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.

Рис. 11-51. Электронная микрофотография жгутика зеленой водоросли Chlamydomonas (поперечный срез) (с любезного разрешения Lewis Tilney). Видна характерная структура типа 9 + 2, свойственная почти всем ресничкам и жгутикам эукариот. Схема, показывающая основные компоненты этой структуры, представлена на рис. 11-53.
по всей длине аксонемы, которая обычно составляет около 10 мкм, в некоторых клетках может достигать 200 мкм.
Если микротрубочки центральной пары одинаковы, и каждая представляет собой отдельный полный цилиндр, то наружные дублеты состоят из двух слившихся микротрубочек - одной полной и одной неполной (субфибриллы А и В соответственно). По линии контакта эти
субфибриллы имеют участок общей стенки. На поперечных срезах видно, что полная микротрубочка дублета образована кольцом из 13
субъединиц, а неполная (субфибрилла В)-только из 11.
11.3.3. Микротрубочки - полые цилиндры, образованные молекулами тубулина [31]
Микротрубочки состоят из молекул тубулина, каждая из которых представляет собой гетеродимер, образованный двумя прочно связанными глобулярными субъединицами. Эти субъединицы - родственные белки (около 450 аминокислот в каждом), получившие название α- и β -
тубули нов. Хотя тубулин присутствует практически во всех клетках эукариот, главным источником его для биохимических исследований служит головной мозг позвоночных: 10-20% растворимого белка, экстрагируемого из мозга большинством методов, составляет тубулин; и это неудивительно, так как отражает высокое содержание микротрубочек в длинных аксонах и в дендритах нервных клеток.
В ходе сборки микротрубочек молекулы тубулина образуют линейные протофиламенты, в которых α -тубулин одного димера контактирует с β-тубулином следующего. Целая микротрубочка содержит 13 таких протофиламентов, уложенных параллельно бок о бок вокруг центральной области, которая на электронных микрофотографиях кажется пустой (рис. 11-52). Так как все протофиламенты уложены параллельно и имеют одинаковую ориентацию, микротрубочки, подобно актиновым филаментам, являются полярными структурами, у которых есть плюс-
концы, растущие быстро, и минус-концы, растущие медленно (см. схему 11-2). Плюс-концы микротрубочек находятся на кончике реснички.
Подобно актину и многим другим белкам цитоскелета, тубулин у большинства организмов закодирован целым семейством близко родственных генов. У одноклеточной зеленой водоросли Chlamydomonas оказалось два гена α-тубулина и два гена β -тубулина, а у плодовой мушки Drosophila-no четыре и того и другого. Аминокислотные по-
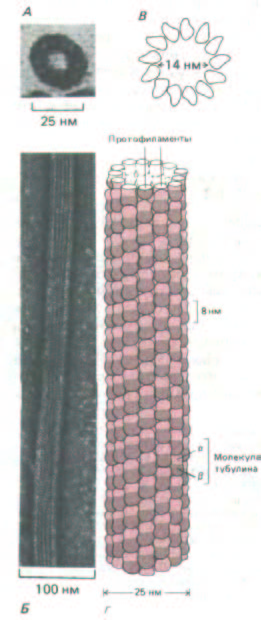
Рис. 11-52. А. Электронная микрофотография поперечного среза микротрубочки, где видно кольцо из 13 субъединиц, каждая из которых соответствует отдельной молекуле тубулина. Б. Электронная микрофотография микротрубочки (негативный контраст). В и Г. Схемы строения микротрубочки, показывающие, как молекулы тубулина образуют стенку цилиндра. В-13 молекул на поперечном разрезе микротрубочки; Г - вид сбоку короткого отрезка микротрубочки с уложенными в продольные ряды (протофиламенты) молекулами тубулина. Каждый из 13
протофиламентов состоит из цепи молекул тубулина, представляющих собой αβ-гетеродимеры. Микротрубочка-полярная структура; разные концы молекулы тубулина (α и β) обращены к разным концам микротрубочки. [С любезного разрешения Richard Linck (А) и Rolley Williams (Б); Г-по данным Linda Amos.]
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
следовательности тубулинов у разных организмов также очень сходны, хотя они не столь консервативны, как у актина: β-тубулин дрожжей, например, на 70% идентичен с куриным β-тубулином, тогда как актины этих видов имеют более 90% идентичных аминокислотных остатков.
Необычайная эволюционная консервативность актина и тубулина может, по крайней мере отчасти, быть следствием структурных ограничений, которые накладываются связыванием их с многочисленными (и разнообразными) белками. Молекулы тубулина, так же как и актина, взаимодействуют не только между собой, но и со многими вспомогательными белками. Как мы увидим, эти белки модифицируют свойства микротрубочек и соединяют их с другими структурами клетки. По-видимому, большинство случайных мутационных изменений нарушают хотя бы одну из функций микротрубочек или актиновых филаментов и поэтому оказываются вредными для организма.
Все ныне известные тубулины, будучи смешаны in vitro, образуют одинаковые микротрубочки. Тем не менее кажется вероятным, что некоторые вариации в структуре тубулинов имеют для клетки функциональное значение. В частности, у высших позвоночных участки тубулинов обоих типов (α и β), содержащие необычно много кислых аминокислотных остатков, обнаруживают явные тканеспецифические различия. Эта область в молекулах тубулина, как полагают, участвует в связывании вспомогательных белков, и изменения ее аминокислотной последовательности могут изменять функции микротрубочек, влияя на связывание этих белков.
11.3.4. Вдоль стенки дублета микротрубочек проходит длинная тонкая нить [32]
Хотя большинство микротрубочек состоит, по-видимому, только из субъединиц тубулина, для построения специальных видов микротрубочек (каковы, например, дублеты микротрубочек в ресничке) используются дополнительные белки. Если заставить микротрубочки ресничек или жгутиков диссоциировать в разбавленном солевом растворе, то из такой смеси удается выделить особенно устойчивые фрагменты субфибриллы А-ленты, состоящие из двух-четырех протофиламентов. Помимо тубулина эти фрагменты содержат белок тектин, образующий длинные нити толщиной 2-3 нм, видимо, родственные промежуточным филаментам. Тектиновые филаменты вытянуты вдоль стенки дублета микротрубочек и, вероятно, способствуют образованию общей стенки А- и В-субфибрилл. Как полагают, эти филаменты или какие-то еще не известные нитевидные молекулы определяют расположение на микротрубочках специальных периодических структур, которые будут описаны ниже.
11.3.5. Аксонема ресничек и жгутиков содержит белковые связки, «ручки» и «спицы» [33]
В аксонеме с микротрубочками связано много других белковых структур, взаимодействие которых обеспечивает ресничку энергией и дает возможность использовать эту энергию для волнообразных движений. Пожалуй, самые важные из этих структур - короткие боковые выступы («ручки»), отходящие от каждого дублета микротрубочек внешнего кольца по направлению к соседнему дублету (рис. 11-53). Пары таких выступов располагаются по всей длине субфибриллы А с интервалами в 24 нм. Они состоят из белка, называемого динеином, и, как мы увидим, играют важную роль в движении ресничек и жгутиков. Другой белок – нексин - образует между соседними дублетами поперечные связи, рас-
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.

Рис. 11-53. Схема поперечного среза реснички (соответствующая микрофотографии на рис. 11-51). Вдоль всей реснички с определенной периодичностью расположены различные структуры, отходящие от микротрубочек (см. табл. 11-2).
положенные на несколько большем расстоянии друг от друга, чем динеиновые «ручки»; по-видимому, они весьма эластичны и стягивают аксонему по окружности, ограничивая скольжение соседних микротрубочек.
От каждого наружного дублета внутрь отходит радиальная «спица», доходящая до внутреннего чехла, окружающего центральную пару одиночных микротрубочек (рис. 11-53). Если смотреть на аксонему сбоку, то все эти структуры - динеиновые ручки, нексиновые связки, радиальные «спицы» и отростки центрального чехла - предстанут в виде боковых выступов, повторяющихся с характерной для каждой из этих структур периодичностью (табл. 11-2).
| Таблица 11-2. Основные белковые структуры аксонемы | |
| Компоненты аксонемы (периодичность расположения вдоль аксонемы) | Функция |
| Тубулин (8 нм) Динеиновые ручки (24 нм) | Главный компонент микротрубочек Выступают из дублетов микротрубочек, взаимодействуют с соседними дублетами, вызывая изгибание |
| Нексиновые связки (86 нм) | Удерживают соседние дублеты микротрубочек вместе |
| Радиальные спицы (29 нм) | Тянутся от каждого из 9 наружных дублетов по направлению к центральной паре |
| Выступы внутреннего чехла (14 нм) | Отходят в виде ряда боковых ручек от центральной пары микротрубочек; вместе с радиальными спицами регулируют характер биения реснички |
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.

Рис. 11-54. Электронная микрофотография изолированной аксонемы (из реснички Tetrahymena), подвергнутой кратковременному воздействию трипсина с целью частично разрушить белковые связи, поддерживающие ее нормальную структуру. После обработки АТР отдельные дублеты микротрубочек скользят относительно друг друга, что ведет к значительному-вплоть до 9-кратного-увеличению длины аксонемы. (F.D.
Warner, D.R. Mitchell, J. Cell Biol. 89: 35-44, 1981. С разрешения Rockefeller University Press.)
|
|
|
|
|
Дата добавления: 2015-08-31; Просмотров: 2540; Нарушение авторских прав?; Мы поможем в написании вашей работы!