КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Свойства цитоплазматических микротрубочек изменяются под влиянием белков, которые с ними связаны
|
|
|
|
11-25
11.4.5. Динамическая нестабильность микротрубочек может служить одной из основ клеточного морфогенеза [45]
В животных клетках цитоплазматические микротрубочки расходятся во всех направлениях от центросомы, к которой прикреплены их минус-концы. Однако большинство животных клеток обладает полярностью. и сборка молекул тубулина в них регулируется таким образом, что многие микротрубочки направляются к определенным участкам клетки. Еще не вполне ясно, как это достигается, но кажется вероятным, что механизм этого процесса основан на динамической нестабильности микротрубочек.
Как мы уже видели, in vitro отдельные микротрубочки чаще всего находятся в одном из двух состояний: в состоянии постепенного роста или же быстрой, «катастрофической» деполимеризации. В клетке они тоже, по-видимому, существуют в этих двух состояниях. Например, в интерфазных фибробластах в культуре среднее время жизни микротрубочи весьма мало, менее 10 мин. Это означает, что весь массив расходящихся от центросомы микротрубочек непрерывно обновляется - одни микротрубочки деполимеризуются, но вместо них вырастают новые.
Присущая микротрубочкам нестабильность позволяет объяснить, как может регулироваться их рост в определенных нужных направлениях в ползущей клетке, например, к переднему краю, а в делящейся к конденсированным хромосомам (разд. 13.5.4). Микротрубочка, у которой оба конца «открыты», в цитоплазме быстро исчезает. Однако центр организации все время порождает новые трубочки. Они направлены случайным образом, а их минус-концы защищены от деполимеризации,

Рис. 11-68. Большой и необычайно упорядоченный центр организации микротрубочек в области ротовой корзинки (цитофаринкса)
инфузории Nassula. Микротрубочки растут регулярным гексагональным пучком от поверхности плоского трехслойного листка, образующего один из элементов сложного ротового аппарата этой клетки. (J. В. Tucker, J. Cell Sci. 6: 385-429, 1970.)
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
так как заякорены в центре организации. Растущая из такого центра микротрубочка может стать стабильной при условии, что ее плюс-конец окажется каким-то образом закрыт - «кэпирован» - и поэтому не будет распадаться. Если микротрубочку кэпировала структура, находящаяся в определенном участке клетки, то между этой структурой и центром организации микротрубочек образуется довольно устойчивая связь. Таким образом, микротрубочки, берущие начало в своем центре организации, могут избирательно стабилизироваться событиями, происходящими в других участках клетки. Как полагают, именно таким путем какие-то не известные пока структуры или факторы, локализованные в определенных областях клеточного кортекса и «захватывающие» плюс-концы микротрубочек, влияют на полярность клеток. Аналогичным образом могла бы определяться и плоскость деления клетки (разд. 13.5.13).
Во многих клетках первоначальная стабилизация микротрубочек путем кэпирования их плюс-концов в дальнейшем закрепляется другими механизмами, обеспечивающими более устойчивую поляризацию клетки. К рассмотрению этих механизмов мы сейчас и перейдем.
11.4.6. Микротрубочки постепенно «созревают» благодаря посттрансляционным модификациям, которым
подвергаются их тубулиновые субъединицы [46]
Непрерывное образование и исчезновение микротрубочек характерно: для клеток, претерпевающих значительную внутреннюю реорганизацию, например делящихся или ползущих по субстрату. Если же клетки становятся частью сформировавшейся ткани, их микротрубочки превращаются в относительно постоянные структуры, особенно в таких клетках, которые, дифференцировавшись, больше уже не делятся (например, нейронах). Это своеобразное «созревание» микротрубочек отчасти зависит от посттрансляционной модификации молекул тубулина и отчасти - от взаимодействия со специфическими белками, ассоциированными с микротрубочками.
Ряд ферментов модифицирует определенные аминокислоты в тубулине. Один из них - тубулин-ацетилтрансфераза, ацетилирующая один из лизиновых остатков α-тубулина. У Chlamydomonas этот фермент находится главным образом в аксонеме жгутика; по-видимому, он ацетилирует молекулы тубулина после их присоединения к дистальному концу аксонемы (разд. 11.3.2). Специфическая дезацетилаза содержится в цитоплазме и удаляет ацетильные группы с неполимерного тубулина. В результате такой локализации этих двух ферментов у Chlamydomonas молекулы тубулина в микротрубочках аксонемы ацетилированы, а в цитоплазматических микротрубочках, большая часть которых быстро обновляется, в основном неацетилированы. Ниже мы еще коснемся вопроса о возможных последствиях ацетилирования тубулина.
Второй, менее обычный способ модификации тубулина - это удаление С-концевого тирозина у молекул α-тубулина, включенных в микротрубочку. Отщепление тирозина катализируется специальным ферментом, имеющимся в цитоплазме многих клеток позвоночных. Как и в случае ацетилирования, есть и фермент, осуществляющий обратную реакцию-восстановление тирозина на С-конце деполимеризованных молекул тубулина. В клетках с быстро обновляющимися микротрубочками большинство молекул тубулина находится в их составе в исходной форме (с тирозином), так как просто не успевают детирозилироваться до отделения от полимера. Зато все более «старые» микротрубочки-те, что «сумели сохраниться» за короткое время обновления,-оказываются обогащены детирозилированным тубулином. Таким образом, и ацети-
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
лирование, и детирозилирование служат маркерами превращения микротрубочек из временно стабилизированной формы в гораздо более устойчивую (рис. 11-69). Если культуру фибробластов обработать препаратом, вызывающим деполимеризацию микротрубочек (разд, 11.4.1), то окажется, что немногие сохранившиеся микротрубочки способны избирательно связывать антитела, узнающие ацетилированную или детирозилированную форму тубулина.
Микротрубочки, образующиеся из ацетилированного и детирозили-рованного тубулина in vitro, ничуть не более устойчивы, чем образованные из обычного, немодифицированного тубулина; значит, модификации, видимо, служат лишь «сигналом» для связывания специфических белков, которые уже стабилизируют микротрубочки и изменяют их свойства в клетках.
[47]
Посттрансляционные модификации тубулина, во-первых, маркируют микротрубочки как «зрелые» и, во-вторых, способствуют их стабилизации. Однако наиболее значительные и важные модификации, которым подвергаются микротрубочки, обусловлены присоединением к ним других белков. Эти- белки, ассоциированные с микротрубочками (БАМ, или MAPs-microtubule-associated proteins), повышают устойчивость микротрубочек к деполимеризации и обеспечивают взаимодействие их с другими компонентами клетки. Зная, сколь разнообразны в клетках функции микротрубочек, можно ожидать, что должно существовать много разных видов БАМ.
Две важные группы БАМ, которые можно выделить из мозга вместе с микротрубочками, - это высокомолекулярные белки (с мол. массой около 200-300 тыс. и даже больше) и may-белки (с мол. массой 40-60 тыс.). Белки обеих групп имеют два домена, один из которых ответствен за связывание с микротрубочками; так как этот домен соединяется одновременно с несколькими неполимерными молекулами тубулина, БАМ ускоряют нуклеацию микротрубочек in vitro. Другой домен, как полагают, обеспечивает взаимодействие микротрубочек с другими клеточными компонентами (рис. 11-70). И высокомолекулярные БАМ, и тау-белки «сидят» на микротрубочках цитоплазмы по всей их длине, что показано с помощью антител к этим белкам.
Выделено уже немало белков, которые избирательно связываются с микротрубочками. Функции большинства из них не известны.
Некоторые, вероятно, играют роль структурных компонентов, стабилизируя микротрубочки и обеспечивая постоянную связь их с другими
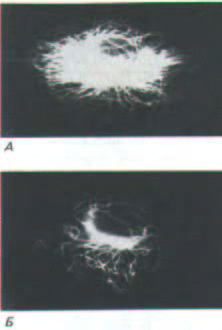
Рис. 11-69. Иммунофлуоресцентные микрофотографии, на которых видно, что одни микротрубочки в цитоплазме культивируемой клетки весьма динамичны (А), а другие относительно стабильны (В). В клетку инъецировали тубулин, ковалентне связанный с небольшой молекулой -
биотином; через час клетку окрашивали сначала меченными флуоресцеином антителами к биотину, а затем антителами к тубулину, меченными другим флуоресцентным красителем - родамином. Быстро обновлявшиеся микротрубочки включали биотинилированный тубулин и поэтому выявлялись антителами к биотину (А), которые покрывали микротрубочки и препятствовали связыванию антител к тубулину. В стабильные микротрубочки биотинилированный тубулин не включался, поэтому они не окрашивались антителами к биотину, но зато красились антителами к тубулину (Б). Стабильные микротрубочки окрашиваются также антителами, узнающими детирозилированный или ацетилированный тубулин. (С любезного разрешения Eric Schulze и Marc Kirschner.)

Рис. 11-70. Регулярно расположенные «боковые ручки», образованные на микротрубочке крупным ассоциированным с микротрубочками белком (БАМ-2) из мозга позвоночных. На электронной микрофотографии показан участок микротрубочки, с которым связано много молекул БАМ-2. Часть молекулы БАМ-2 отходит от микротрубочки в сторону, как показано на схеме внизу (Фото любезно предоставили William Voter и Harold Erickson.)
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
компонентами клетки (в том числе с другими элементами цитоскелета и мембранами некоторых органелл); другие же ответственны за перемещение органелл вдоль по микротрубочкам.
|
|
|
|
|
Дата добавления: 2015-08-31; Просмотров: 869; Нарушение авторских прав?; Мы поможем в написании вашей работы!