КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Организация цитоскелета
|
|
|
|
11-29
11.5.4. Сборка промежуточных филаментов может контролироваться с помощью фосфорилирования [55]
Изолированные промежуточные филаменты (ПФ) в ионной среде, соответствующей цитоплазме, чрезвычайно стабильны; более того, сколько-нибудь значительного пула неполимеризованных белков ПФ (какой
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
имеется в случае актина и тубулина) в клетке нет. И все же клетка явно может регулировать число, длину и расположение своих промежуточных филаментов, что указывает на ее способность контролировать их сборку и разборку. Важный фактор этого контроля-фосфорилирование определенных остатков в белках ПФ. Виментин, например, существует как в нефосфорилированной, так и в фосфорилированной форме. Если фосфорилировать изолированные виментиновые нити с помощью протеинкиназы, они распадаются на меньшие фрагменты. Однако самый впечатляющий пример того, насколько важную роль играет в контроле разборки ПФ фосфорилирование, - это ядерные ламины, которые подвергаются деполимеризации всякий раз, когда клетка вступает в митоз, 11.5.5. Ядерная ламина образована особым классом промежуточных филаментов [56]
Ядерная ламина - это белковая сеть (обычно толщиной от 10 до 20 нм), подстилающая изнутри поверхность внутренней ядерной мембраны (см. рис. 9-1). Она представляет собой прямоугольную решетку из промежуточных филаментов (рис. 11-76, Д), построенных у млекопитающих из трех белков ПФ типа Vl-ламинов А, В и С (см. рис. 11-74 и табл. 11-3), Ламины образуют димеры, у которых имеется стержневидный домен и две глобулярные головки на одном из концов (рис. 11-76, Б). При подходящих рН и ионной силе димеры самопроизвольно ассоциируют, образуя филаменты, которые по толщине и повторяющейся структуре сходны с цитоплазматическими ПФ.
Однако по ряду признаков ядерные ламины отличаются от белков ПФ цитоплазмы. Наиболее очевидное отличие-это организация образуемых филаментов в прямоугольную решетку (рис. 11-76, А), хотя для такой организации, видимо, необходимо объединение их с другими белками. Кроме того, ядерная ламина - структура очень динамичная. Когда клетки млекопитающих вступают в митоз, кратковременное фосфорилирование нескольких остатков серина в ламинах вызывает обратимую диссоциацию ядерной ламины на тетрамеры гиперфосфорилированных ламинов А и С и связанного с мембраной ламина В. При возвращении клетки в интерфазу ламины дефосфорилируются, и вокруг разошедшихся хромосом вновь образуется цельная ядерная оболочка (разд. 13.5.11).
11.5.6. Кератиновые филаменты удивительно разнообразны [52]
Из всех типов промежуточных филаментов наиболее стабильные и долгоживущие - кератиновые, они же и самые разнообразные.
Эпителии с примитивной организацией, например в развивающемся эмбрионе, а также некоторые зрелые ткани (такие, как печень) содержат кератины двух типов - один кислый и один нейтральный. В эпителиях других органов (например, языка, мочевого пузыря, потовых желез) имеются шесть или больше различных кератинов, причем их конкретный спектр зависит от анатомической локализации. Кератиновые филаменты ввиду их многообразия и стабильности могут служить своего рода «отпечатками пальцев», позволяющими уточнить происхождение некоторых опухолей эпителиальной природы.
Еще разнообразнее кератины в эпидермисе, который представляет собой плотный многослойный эпителий (разд. 17.4.2). В клетках разных слоев эпидермиса экспрессируются разные наборы кератинов. Кератиновые филаменты в них постепенно сшиваются поперечными ковалентными связями друг с другом и с ассоциированными белками, и по мере
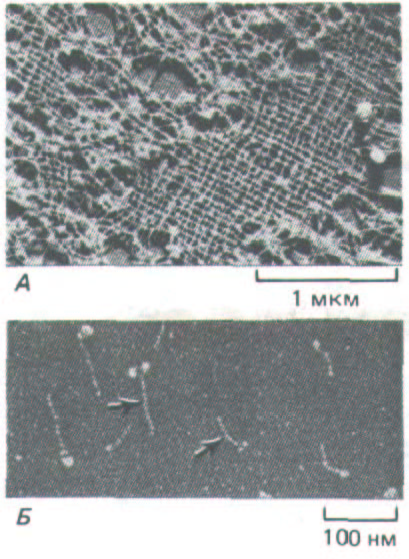
Рис. 11-76. А. Электронная микрофотография участка ядерной ламины в ооците Xenopus (препарат получен методом лиофилизации и напыления металлом). Ламину образует высокоупорядоченная прямоугольная сеть из промежуточных филаментов, состоящих из ядерных ламинов.
Б. Изолированные димеры ламина (L) (электронная микрофотография, напыление металлом). По форме они напоминают мышечный миозин (М): у них есть стержневидный хвост и две глобулярные головки, но они гораздо меньше. Глобулярные головки образованы двумя большими С-
концевыми доменами. (С любезного разрешения Ueli Aebi.)
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
гибели клеток в самых наружных слоях эпидермиса поперечносшитый кератиновый скелет становится важнейшим защитным барьером на поверхности тела. Специализированные эпителиальные клетки, образующие такие поверхностные структуры, как волосы, когти и перья, обеспечивают дополнительные локальные вариации в наборе кератинов. Таким образом, промежуточные филаменты защищают животное от потери тепла и воды, предоставляют ему «оружие» и средства камуфляжа или, наоборот, привлечения партнера (окраска).
11.5.7. Какова функция промежуточных филаментов?
Животные клетки могут обходиться и без промежуточных филаментов. В ЦНС глиальные клетки, вырабатывающие миелин, совершенно лишены их. Если культивируемым фибробластам сделать внутриклеточную инъекцию антител к белкам промежуточных филаментов, то последние разрушатся, и это не окажет заметного влияния на организацию или поведение клеток. Кажется вероятным, что главная функция большинства ПФ состоит в чисто механической «поддержке» клетки и ее ядра. Промежуточные филаменты в эпидермальных пластах образуют «трансклеточную»
(соединяющую множество клеток воедино) сеть, роль которой, скорее всего, заключается в противодействии внешним нагрузкам. Нейрофиламенты в нервных волокнах противостоят механическим деформациям, возникающим при движении животного, иначе эти длинные тонкие цилиндры из цитоплазмы легко рвались бы. Десминовые нити в клетках мышц создают механическую опору для саркомеров, а виментиновые окружают (и, вероятно, поддерживают) крупные жировые капли в жировых клетках.
Но если функция промежуточных филаментов сводится всего лишь к сопротивлению растягивающим силам, для чего нужно так много различных вариантов их белковых субъединиц? Какова роль вариабельных частей их молекул - ведь они как будто бы не участвуют в построении самого филамента? Детальных ответов на эти вопросы пока нет, но ясно, что как характер опорной функции промежуточных филаментов, так и способ их соединения с другими компонентами клетки сильно различаются в клетках разного типа. Например, десминовые филаменты, скрепляющие края Z-дисков в поперечнополосатых мышцах, по-видимому, имеют участки для связывания специфических белков Z-диска;
нейрофиламенты подвергаются меньшим нагрузкам, но они могут соединяться вместе боковыми поверхностями, образуя непрерывный «трос» до метра и больше длиной; вероятно, именно поэтому

Рис. 11-77. Электронная микрофотография промежуточных филаментов двух типов, встречающихся в нервной ткани (препарат после быстрого замораживания и глубокого травления). А. Нейрофиламенты в аксоне соединены многочисленными поперечными белковыми сшивками;
как полагают, такая организация придает этому длинному клеточному отростку большую прочность на разрыв. По-видимому, сшивки образованы длинными неспиральными участками С-концевой части наиболее крупного белка нейрофиламентов (см. рис. 11-74). Б. Промежуточные филаменты (называемые глиальными филаментами) в астроците. Они подвергаются меньшим механическим нагрузкам. Их поверхность довольно гладкая, и поперечных сшивок между ними мало. (С любезного разрешения N. Hirokawa.)
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
нейрофиламенты имеют по всей своей длине большие выступы, которых нет у других промежуточных филаментов (рис. 11-77).
Различные потенции к связыванию других белков могут обеспечиваться вариабельными участками белков промежуточных филаментов, Влияя на свойства филамента, эти вариабельные участки определяют не только его способность к самосборке, но и то, как он будет взаимодействовать с другими компонентами клетки (например, с микротрубочками и плазматической мембраной). Это совершенно иная стратегии чем в случае двух других важнейших элементов цитоскелета - актиновных филаментов и микротрубочек; как мы уже знаем, эти полимеры в основном инвариантны по структуре, а к выполнению различных функций они приспосабливаются с помощью разных наборов актин-связывающих белков и белков, ассоциированных с микротрубочками. Таким образом, роль вариабельных участков в белках промежуточных филаментов та же, что и у вспомогательных белков актиновых филаментов и микро-трубочек, - разница лишь в том, что одни ковалентно связаны с субъединицами филамента, а другие представляют собой отдельные молекулы.
Заключение
Промежуточные филаменты (ПФ) - это полимеры, по структуре подобные канатам, собранным из нитевидных полипептидов. По-
видимому, они поддерживают структуру клеток или противостоят растягивающим нагрузкам. Существует много тканеспецифических форм
ПФ, построенных из различных полипептидов: кератиновые филаменты эпителиальных клеток, нейрофиламенты нейронов, глиальные
филаменты астроцитов и шванновских клеток, десминовые филаменты мышечньа волокон и виментиновые филаменты фибробластов и клеток
многих других типов. Отдельное семейство белков ПФ составляют ядерные ламины, из которых построена волокнистая пленка (ламина), выстилающая изнутри оболочку ядра; они имеются во всех эукариотических клетках.
Полипептиды, входящие в состав промежуточных филаментов различных типов, различаются по аминокислотной
последовательности, а также - и очень сильно - по молекулярной массе. Однако у всех имеется гомологичный центральный домен, который при
димеризации белка образует жесткую структуру из обвивающих друг друга спиралей. Такие димерные субъединицы складываются в большие
пучки «внахлест», формируя промежуточные филаменты. Стержневидные домены субъединиц при этом создают структурную сердцевину ПФ, а
глобулярные домены на обоих концах выступают наружу и обусловливают разнообразие свойств ПФ. Именно благодаря этой вариабельности
механические свойства ПФ и взаимодействия их с другими клеточными компонентами приспособлены к специфическим нуждам клеток того или
иного типа.
До сих пор мы рассматривали микротрубочки, актиновые филаменты и промежуточные филаменты так, как будто это независимые составные части цитоскелета. В действительности, конечно, различные элементы цитоскелета должны быть связаны в единое целое, а их функции скоординированы, чтобы клетка могла осуществлять разного рода движения и изменять свою форму. Например, когда находящийся в культуре фибробласт округляется, готовясь к делению, реорганизуется весь его цитоскелет в целом: исчезают стрессовые волокна и цитоплазматические микротрубочки, появляется митотическое веретено,
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
а затем и сократимое кольцо, и все это происходит как единая контролируемая цепь событий.
В этом разделе мы рассмотрим взаимодействия между главными системами филаментов цитоскелета в связи с тремя его функциями.
Сначала мы выясним, каким образом цитоскелет организует содержимое цитоплазмы, включая те ее компоненты, которые обычно считают свободно растворенными. Затем мы перейдем к вопросу о том, как координированная работа цитоскелета обеспечивает направленное передвижение животной клетки по твердому субстрату. И наконец, мы обсудим, как цитоскелет порождает те многочисленные морфологические изменения, которые происходят в процессе развития зародыша. В результате этого рассмотрения станет ясно, насколько фрагментарны пока наши представления о молекулярных механизмах этих фундаментальных процессов.
11.6.1. В цитоплазме имеется сложная трехмерная сеть белковых нитей [58]
Мы уже знаем, что кортикальная цитоплазма многих животных клеток содержит сети из поперечносшитых актиновых филаментов.
Подобные же сети, но образованные взаимодействующими актиновыми филаментами, микротрубочками и промежуточными филаментами, пронизывают всю цитоплазму. Наиболее отчетливо они видны после экстракции клеток неионным детергентом, которая удаляет фосфолипиды и растворимые белки. Если обработанные таким образом клетки быстро заморозить и подвергнуть глубокому травлению, то металлическая реплика с полученного препарата выявит поразительную картину цитоскелета (рис. 11-78). Различные типы белковых филаментов можно распознать по их толщине, а в некоторых случаях и по расположению их белковых субъединиц. Часто бывает видно, что соседние филаменты соединены более тонкими нитями, белки которых тоже были в ряде случаев идентифицированы с помощью антител. Это оказались различные виды белков, ассоциированных с микротрубочками, и длинные гибкие боковые отростки, отходящие от субъединиц промежуточных филаментов некоторых типов (см. рис. 11-77, А). Однако в большинстве случаев белки, образующие поперечные сшивки, пока не известны.
В клетках, не обработанных детергентом, структура цитоплазмы еще сложнее. Пространство между филаментами цитоскелета заполнено зернистым «основным веществом» ("ground substance"), которое, как считают, представляет собой очень концентрированную смесь «растворимых»
белков, имеющихся в живой клетке. Разнообразные мембранные органеллы тоже погружены в этот плотный матрикс и соединены с филаментами цитоскелета тонкими белковыми нитями. И гранулярного материала, и органелл тем больше, чем ближе к центральной области клетки, где сосредоточены микротрубочки и промежуточные филаменты и где, как можно увидеть при помощи светового микроскопа с видеоприставкой, происходит большая часть процессов цитоплазматического транспорта. В более периферийных участках значительно гуще сеть актиновых филаментов, которые как бы вытесняют оттуда большую часть мембранных органелл, а возможно, и какую-то долю гранулярного материала (рис.
11-79). Эта густая сеть прикреплена к плазматической мембране и соответствует богатому актином кортексу клетки, о котором уже говорилось ранее (разд. 11.2).
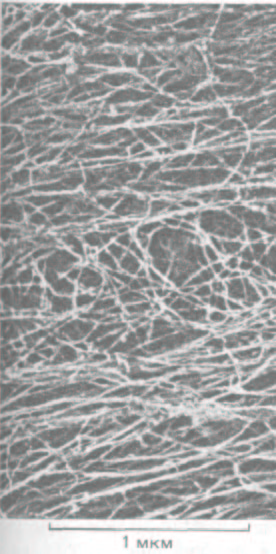
Рис. 11-78. Цитоплазма фибробласта, экстрагированного неионным детергентом (электронная микрофотография, глубокое травление).
Большинство тянущихся слева направо прямых волокон, образующих рыхлые пучки, - это актиновые филаменты, а перепутанные волокна в середине фото - в основном промежуточные филаменты. (С любезного разрешения John Heuser и Marc Kirschner.)
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
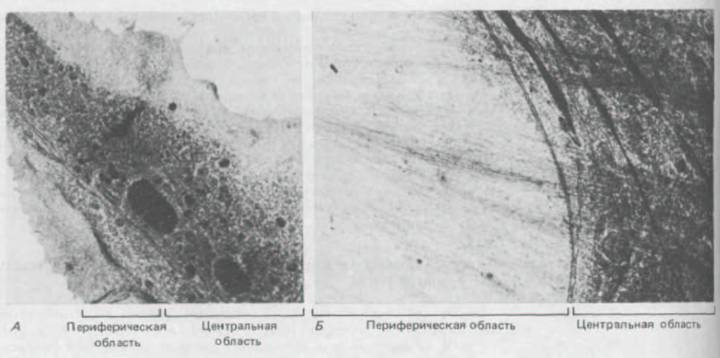
Рис. 11-79. Трансмиссионная электронная микрофотография фибробласта, замороженного «живьем», выявляет на периферии клетки участок, свободный от органелл, а ближе к центру-наоборот, богатую органеллами область (А). Граница между этими двумя областями при сильном увеличении видна более отчетливо (Б). (Р. С. Bridgman, В. Kachar, Т. S. Reese, J. Cell Biol. 102: 1510-1521, 1986. С разрешения Rockefeller Univ.
Press.)
11.6.2. Насколько упорядочена организация цитоплазмы? [59]
Когда мембранные органеллы цитоплазмы быстро перемещаются с места на место, они движутся по белковым «дорожкам», с которыми соединены специальными мостиками. Как мы уже говорили, движение вдоль микротрубочек осуществляется с помощью кинезина и динеино-
подобных белков (разд. 11.4.9), а вдоль актиновых филаментов с помощью миозиноподобных белков (разд. 11.2.4). Кластеры рибосом в цитозоле тоже нередко находятся в ассоциации с филаментами; при экстрагировании клеток неионными детергентами значительная часть аппарата белкового синтеза остается связанной с цитоскелетом. Даже растворимые ферменты, в том числе некоторые ферменты гликолиза, по-видимому, «сидят» на специфических участках миофибрилл в мышечных клетках и стрессовых волокон в фибробластах, и здесь их можно выявить с помощью флуоресцентных антител.
Степень структурной организованности цитозоля в настоящее время является предметом споров. Источник наших знаний об идущих в цитозоле процессах - это в основном биохимические исследования, а они начинаются с гомогенизации клетки, так как только после этого можно определять активность ферментов и выделять их в очищенном виде. Успехи такого подхода привели многих биохимиков к представлению о цитозоле как о простом растворе ферментов. Однако другие исследователи склонны полагать, что едва ли не большинство ферментов цитозоля собрано в группы по принадлежности к тому или иному биохимическому пути и прикреплено к цитоскелету так, чтобы обеспечить более быстрое и эффективное прохождение метаболитов по каждому из таких путей. Поскольку это прикрепление, вероятно, непрочно и связи легко разрушаются, для его убедительной демонстрации могут потребоваться новые методы - такие, например, как инъекции в живые клетки белков с флуоресцентными метками (разд. 4.2.3).
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
|
|
|
|
|
Дата добавления: 2015-08-31; Просмотров: 1743; Нарушение авторских прав?; Мы поможем в написании вашей работы!