КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Базальная мембрана-это специализированная форма внеклеточного матрикса, содержащая в основном
|
|
|
|
14-14
14.2.13. Фибронектин внеклеточный гликопротеин, способствующий адгезии между клеткой и матриксом [21]
Внеклеточный матрикс содержит несколько адгезивных гликопротеинов, которые связываются и с клетками, и с другими макромолекулами матрикса, способствуя прикреплению клеток к матриксу. Из них лучше всего охарактеризован фибронектнн - крупный, образующий фибриллы гликопротеин, встречающийся во всем животном царстве. Это димер, состоящий из двух одинаковых субъединиц (каждая почти из 2500 аминокислотных остатков); субъединицы соединены парой дисульфидных связей вблизи от карбоксильных концов и свернуты в серию глобулярных доменов, разделенных участками гибкой полипептидной цепи (рис. 14-46). Как показывает секвенирование, молекула фибронектина состоит в основном из трех коротких много раз повторяющихся последовательностей аминокислот, и это позволяет предполагать, что ген, кодирующий фибронектин, образовался в результате многократной дупликации трех малых генов.
Существуют три формы фибронектина: 1) растворимая димерная форма (фибронектин плазмы}, которая циркулирует в крови и тканевых жидкостях и, как полагают, способствует свертыванию крови, заживлению ран и фагоцитозу; 2) олигомеры фибронектина, которые могут быть временно прикреплены к поверхности клеток (поверхностный фибронектин); 3) труднорастворимая фибриллярная форма фибронектина во внеклеточном матриксе (матриксный фибронектин). В поверхностных
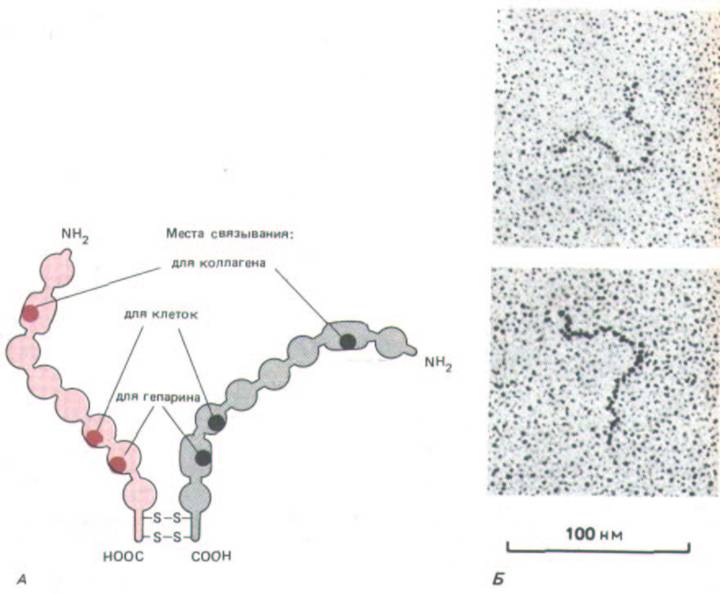
Рис. 14-46. А. Схематическое изображение структуры димера фибронектина. Б. Электронные микрофотографии отдельных молекул, напыленных платиной. Две полипептидные цепи сходны, но не идентичны; вблизи карбоксильного конца они соединены двумя дисульфидными связями. Каждая цепь свернута в ряд глобулярных доменов, соединенных гибкими полипептидными сегментами. Отдельные домены предназначены для связывания с той или иной молекулой или клеткой (это указано для трех доменов). Для простоты отмечены не все известные участки связывания. [Б-J. Engel et al. J. Моl. В.о.. 150: 97-120, 1981. Copyright Academic Press Inc. (London) Ltd.]
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
и матриксных агрегатах димеры фибронектина связаны друг с другом дополнительными дисульфидными сшивками.
Фибронектин - многофункциональная молекула, в которой различные глобулярные домены играют разную роль. Например, один домен связывается с коллагеном, другой - с гепарином, третий - со специфическими рецепторами на поверхности клеток различных типов и т. д. (см. рис.
14-46). Таким образом фибронектин участвует в организации матрикса и способствует прикреплению к нему клеток.
Роль различных доменов, в особенности доменов, связывающихся с клетками, была изучена путем расщепления молекулы на отдельные домены протеолитическими ферментами или путем синтеза специфических белковых фрагментов химическим методом или с помощью рекомбинантных ДНК. Так, из протеолитических фрагментов был выделен домен, ответственный за связывание с клеткой, и определена его аминокислотная последовательность. Были приготовлены синтетические пептиды, соответствующие различным сегментам этого домена, и удалось выяснить, что за связывающую активность ответственна специфическая трипептидная последовательность (Arg-Gly-Asp, или R-G-D). Пептиды, содержащие эту RGD-последовательность, конкурируют за места связывания на клетке и таким образом ингибируют прикрепление клеток к фибронектину; когда же эти пептиды связываются с твердой поверхностью, они обусловливают прикрепление к ней клеток. RGD-
последовательность содержится не только в фибронектине - она является общей для многочисленных внеклеточных адгезивных белков и узнается целым семейством гомологичных рецепторов клеточной поверхности, связывающих эти белки (разд. 14.2.17). Хотя в молекулах, узнаваемых этими рецепторами, имеется общая трипептидная последовательность, каждый рецептор специфически узнаёт свою собственную небольшую группу адгезивных молекул; таким образом, связывание с рецептором должно также зависеть и от других участков адгезивной белковой последовательности.
Фибронектин важен не только для клеточной адгезии, но и для миграции клеток. В зародышах беспозвоночных и позвоночных он, по-
видимому, во многих случаях направляет миграцию. Например, большие количества фибронектина находятся вдоль пути передвижения клеток проспективной мезодермы при гаструляции у амфибий (разд. 16.1.4). Миграцию этих клеток можно подавить либо путем инъекции в бластоцель антител к фибронектину, либо путем введения полипепти-дов, содержащих трипептид, связывающийся с клетками, но без тех доменов фибронектина, которые связываются с матриксом. Полагают, что фибронектин способствует миграции клеток, помогая их прикреплению к матриксу. Такое действие должно быть тонко сбалансировано так, чтобы сцепление клеток с матриксом происходило, но не приводило к их иммобилизации. Позднее мы вернемся к вопросу о том, как может быть достигнут такой баланс многочисленными адгезивными молекулами, которые участвуют в определении путей морфогенетических движений.
14.2.14. Множественные формы фибронектина синтезируются при альтернативном сплайсинге РНК [22]
Фибронектин, будучи представителем обширного семейства RGD-coдержащих адгезивных молекул, и сам по себе, как уже отмечалось, может существовать во многих формах; даже в одном димере полипептидные цепи могут слегка различаться. И тем не менее все различные полипептидные цепи фибронектина кодируются одним большим геном; у крысы его длина составляет более 70 тысяч п. н., и он содержит около 50
экзонов, т. е. это один из самых крупных генов охарактеризованных
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
до настоящего времени. При транскрипции получается одна длинная молекула РНК, которая подвергается сплайсингу в разных вариантах, при которых в зависимости от типа клеток образуется одна или несколько из примерно 20 различных мРНК. Неясно, чем определяется выбор вариантов сплайсинга РНК и каковы функциональные различия получающихся при этом полипептидных цепей. Судя по некоторым данным, одной из функций альтернативного сплайсинга РНК-транскрипта для фибронектина человека состоит в добавлении к определенным молекулам фибронектина дополнительного связывающегося с клетками домена, отличного от RGD-содержащего участка.
Фибронектин - не единственный секретируемый гликопротеин, участвующий в адгезии между клетками и матриксом. Например, тенасцин тоже является внеклеточным адгезивным гликопротеин ом, однако он встречается значительно реже, чем фибронектин, и наиболее обычен в эмбриональных тканях. В нервной системе он секретируется глиальными клетками, и полагают, что некоторые нейроны связываются с ним через посредство специфического протеогликана клеточной поверхности. Тенасцин представляет собой крупный комплекс из шести связанных дисульфидными сшивками полипептидных цепей, расходящихся из центра, подобно спицам колеса (см. рис. 14-51).
Некоторые клетки, особенно эпителиальные, секретируют внеклеточный адгезионный гликопротеин иного типа, называемый ламинином, который является одним из основных белков всех базальних мембран. Он связывается как с эпителиальными клетками (а также с клетками некоторых других типов), так и с коллагеном типа IV-главным типом коллагена в базальной мембране.
коллаген типа IV, протеогликаны и ламинин [23]
Базальная мембрана представляет собой тонкий слой специализированного внеклеточного матрикса, подстилающий пласты эпителиальных клеток; кроме того, она окружает отдельные мышечные волокна, жировые клетки и шванновские клетки (которые, наматываясь на перифери-
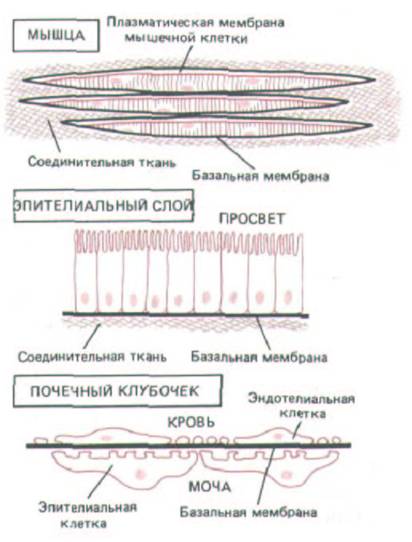
Рис. 14-47. Три варианта расположения базальной мембраны (представлена черной линией). Базальная мембрана клетки может окружать (например, мышечные), подстилать слои эпителиальных клеток или располагаться между двумя клеточными слоями (как в почечных клубочках).
Обратите внимание, что в почечных клубочках в обоих слоях клеток имеются разрывы, так что барьером проницаемости, определяющим, какие молекулы из крови перейдут в мочу, служит базальная мембрана. Поскольку в клубочках эта мембрана-результат слияния двух базальных мембран, образованных эндотелиальными и эпителиальными клетками, то она вдвое толще большинства таких мембран.
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.

Рис. 14-48. Базальная мембрана роговицы куриного эмбриона. Микрофотография, полученная с помощью сканирующего электронного микроскопа. Часть эпителиальных клеток (Эп) была удалена, чтобы показать верхнюю поверхность базальной мембраны (БМ). Обратите внимание, что с нижней поверхностью мембраны взаимодействует густая сеть коллагеновых фибрилл (К). Макромолекулы базальной мембраны синтезируются лежащими на ней эпителиальными клетками. (С любезного разрешения Robert Trelstad.)
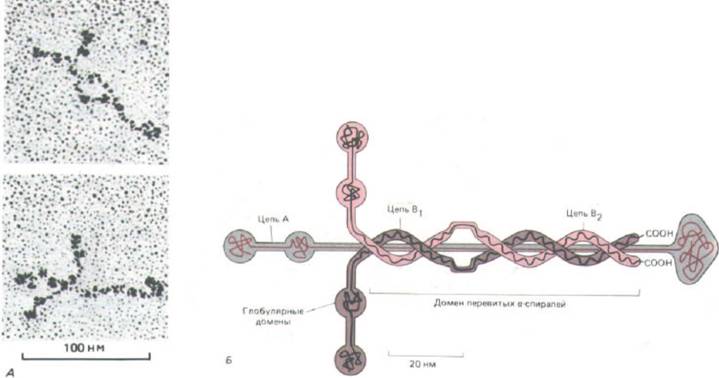
Рис. 14-49. А. Электронные микрофотографии молекул ламинина (препарат, напиленный платиной). Б, Схема структуры ламинина. Этот мультидоменный гликопротеин состоит из трех полипептидных цепей (А, В, и В2), соединенных дисульфидными связями в форме асимметричного креста. [A-J. Engel et al. J. Моl. BioL, 150, 97-120, 1981. Copyright Acad. Press Inc. (London) Ltd; Б-по В.L. Hogan et al. In: Basement Membranes (S.
Shibata, ed.), pp. 147-154. Amsterdam, Elsevier, 1985.]
ческие нервные волокна, образуют миелин). Таким образом, базальная мембрана отделяет эти клетки или клеточные слои от окружающей или подстилающей их соединительной ткани. В других местах, например в почечных клубочках или легочных альвеолах, базальная мембрана расположена между двумя различными слоями клеток и служит здесь высокоэффективным фильтром (рис. 14-47). Однако роль базальных мембран не сводится просто к функциям структурной опоры и фильтра. Они способны определять полярность клеток, влиять на клеточный метаболизм, упорядочивать белки в прилегающих плазматических мембранах, вызывать дифференцировку клеток и, подобно фибронектину, служить специфическими «магистралями» для клеточных миграций.
Базальная мембрана в основном синтезируется лежащими на ней клетками (рис. 14-48). Это, по существу, плотная прослойка из коллагена типа IV (см. рис. 14-42) с дополнительными специфическими молекулами по обе ее стороны, которые способствуют прикреплению ее к соседним клеткам или матриксу. Хотя состав базальных мембран несколько меняется от ткани к ткани и даже от участка к участку (разд. 14.2.17), все эти мембраны содержат коллаген типа IV вместе с протеогликанами (в
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
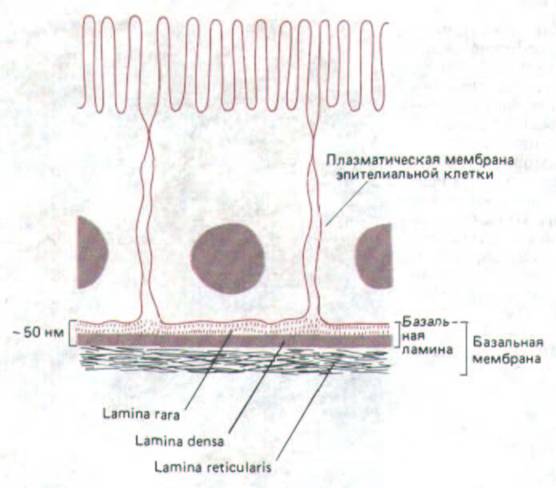
Рис. 14-50. Структура базальной мембраны, подстилающей эпителиальные клетки, как она видна на поперечных срезах в электронном микроскопе.
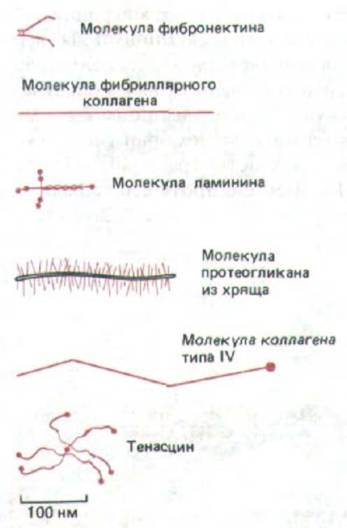
Рис. 14-51. Сравнение формы и размеров некоторых важнейших макромолекул внеклеточного матрикса.
основном гепарансульфатами) и гликопротеинами лиминином и энтактином. Ламинин представляет собой крупный (мол. масса около 850000)
комплекс из трех очень длинных полипептидных цепей, расположенных в форме креста и удерживаемых вместе дисульфидными связями (рис. 14-
49). Подобно фибронектину, он состоит из нескольких функциональных доменов: один из них связывается с коллагеном типа IV, один - с гепарансульфатом и один или несколько других - с белковыми рецепторами ламинина на клеточной поверхности. Полагают также, что с каждой молекулой ламинина прочно связана одна гантелевидная молекула энтактина в месте соединения коротких ветвей «креста» с длинной ветвью.
Как видно на электронно-микроскопических препаратах после обычной фиксации и контрастирования, большинство базальних мембран состоит из двух слоев: электронопрозрачного слоя (lamina lucida, или гага), прилегающего к плазматической мембране клеток, находящихся на базальной мембране (обычно это эпителиальные клетки), и лежащего под ним электроноплотного слоя (lamina densa). В некоторых случаях имеется еще третий слой, содержащий коллагеновые фибриллы (lamina reticularis) и связывающий базальную мембрану с подлежащей соединительной тканью. Некоторые авторы называют «базальной мембраной» весь комплекс из трех слоев (рис. 14-50), толщина которого обычно достаточна для того, чтобы его можно было видеть в обычный микроскоп. Детальная молекулярная организация базальной мембраны еще не исследована, хотя электронная микроскопия с использованием меченых антител показывает, что lamina densa, по-видимому, состоит в основном из коллагена типа IV с молекулами протеогликана на обеих ее сторонах; как полагают, ламинин присутствует главным образом на той стороне lamina densa, которая обращена к плазматической мембране, где он участвует в прикреплении эпителиальных клеток к базальной мембране, тогда как фибронектин помогает связыванию макромолекул матрикса и клеток соединительной ткани на противоположной стороне.
На рис. 14-51 сравниваются размеры и форма некоторых важнейших макромолекул базальной мембраны.
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
14.2.16. Вязальные мембраны выполняют многообразные и сложные функции [24]
Функции базальной мембраны чрезвычайно разнообразны. В почечных клубочках необычно толстая базальная мембрана работает как молекулярный фильтр, регулируя переход молекул из крови в мочу (см. рис. 14-47). Для этой функции, по-видимому, необходимы протеогликаны, так как их удаление с помощью специфических ферментов ведет к утрате фильтрующих свойств мембраны. Базальная мембрана может также служить избирательным барьером для клеток. Например, базальная мембрана, подстилающая эпителиальный слой, обычно предотвращает контакт фибробластов подлежащей соединительной ткани с эпителиальными клетками, но не препятствует прохождению через нее макрофагов, лимфоцитов и нервных волокон.
Базальная мембрана играет важную роль в процессе регенерации ткани после повреждения. При нарушении целости мышечной, нервной или эпителиальной ткани сохранившаяся базальная мембрана служит субстратом для миграции регенерирующих клеток. Таким образом легко восстанавливается исходная архитектура ткани. Наиболее яркий пример роли базальной мембраны в регенерации мы находим при изучении нервно-
мышечного соединения, в котором нервная клетка передает стимул волокну скелетной мышцы.
В месте нервно-мышечного контакта (синапса) базальная мембрана имеет особое химическое строение, распознаваемое, например, антителами, которые связываются с ней исключительно в этом участке. Одна из функций базальной мембраны здесь состоит, по-видимому, в координировании пространственной организации компонентов по обе стороны синапса. Данные в пользу центральной роли базальной мембраны синаптического соединения в восстановлении синапса после повреждения мышцы или нерва будут обсуждаться в гл. 19 (разд. 19.8.3). Такие исследования ясно показывают, что мы еще многого не знаем о химической и функциональной специализации базальной мембраны. Они позволяют также предположить, что минорные (но пока не идентифицированные) компоненты внеклеточного матрикса могут играть решающую роль в управлении процессами морфогенеза в эмбриональном развитии.
14.2.17. Интегрины способствуют связыванию клеток с внеклеточным матриксом [25]
Чтобы понять, как внеклеточный матрикс взаимодействует с клетками, нужно изучить молекулы клеточной поверхности, связывающиеся с компонентами матрикса, а также молекулы самого матрикса. Как уже отмечалось, некоторые протеогликаны являются интегральными компонентами плазматической мембраны; их сердцевинный белок либо пронизывает липидный бислой, либо ковалентне присоединен к нему.
Связываясь с большинством компонентов внеклеточного матрикса, эти протеогликаны способствуют прикреплению клеток к матриксу. Однако компоненты матрикса тоже прикрепляются к клеточной поверхности с помощью специфических рецепторных протеогликанов. Ввиду таких сложных взаимодействий между макромолекулами матрикса во внеклеточном пространстве вопрос о том, где кончаются компоненты плазматической мембраны и где начинается внеклеточный матрикс, - в значительной степени семантический. Например, гликокаликс клетки часто состоит из компонентов обеих этих структур (см. разд. 6.3.1).
Рецепторы матрикса отличаются от поверхностных клеточных рецепторов для гормонов и других растворимых сигнальных молекул тем, что
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
связывают свои лиганды с относительно низким сродством (Ка = 106-108 л/моль), и тем, что их концентрация на поверхности клеток примерно в 10-
100 раз больше. Поэтому такие рецепторы, вероятно, могут функционировать кооперативно, а клетки могут отвечать на организованную группу лигандов в матриксе, а не на отдельные молекулы. В пользу этого предположения говорит то, что растворимые фрагменты матрикса, прикрепляющиеся к клеткам, обычно не вызывают у клетки ответ, вызываемый теми же компонентами, закрепленными в матриксе.
Рецептор фибронектина на фибробластах млекопитающих - один из наиболее изученных рецепторов для компонентов матрикса.
Первоначально он был идентифицирован как гликопротеин плазматической мембраны, который связывается в колонке с фибронектином и может быть элюирован с помощью небольшого белка, содержащего прикрепляющуюся к клетке последовательность RGD (разд. 14.2.13). Рецептор представляет собой нековалентно связанный комплекс из двух различных высокомолекулярных полипептидных цепей, называемых α- и β-цепями.
Он работает как трансмембранный линкер, осуществляя взаимодействие между актином цитоскелета внутри клетки и фибронектином во внеклеточном матриксе (рис. 14-52). Позднее мы увидим, что такие взаимодействия через плазматическую мембрану могут поляризовать и клетку и матрикс. Было охарактеризовано много других рецепторов для матрикса, в том числе таких, которые связывают коллаген и ламинин, и показано, что они родственны рецептору фибронектина на фибробластах. Называемые интегринами, все они являются гетеродимерами с α- и β-цепями, гомологичными цепям рецептора для фибронектина. Вероятно, большинство из них узнаёт последовательности RGD в компонентах матрикса, с которыми они связываются.
Имеется по крайней мере три семейства в огромном суперсемействе интегринов; члены одного семейства имеют одну общую α-цепь, но различаются своими β-цепями. Одно из семейств включает рецептор фибробластов для фибронектина и по меньшей мере еще пять других членов.
Другое семейство включает рецептор на кровяных пластинках, связывающий некоторые компоненты матрикса, в том числе фибронектин и фибриноген - белок, взаимодействующий с пластинками в процессе свертывания крови: при болезни Гланцмана - наследственном дефиците этих рецепторов - у больных плохо свертывается кровь. Третье семейство интегринов составляют рецепторы, находящиеся главным образом на поверхности лейкоцитов; один из них носит название LFA-1 (lympho-
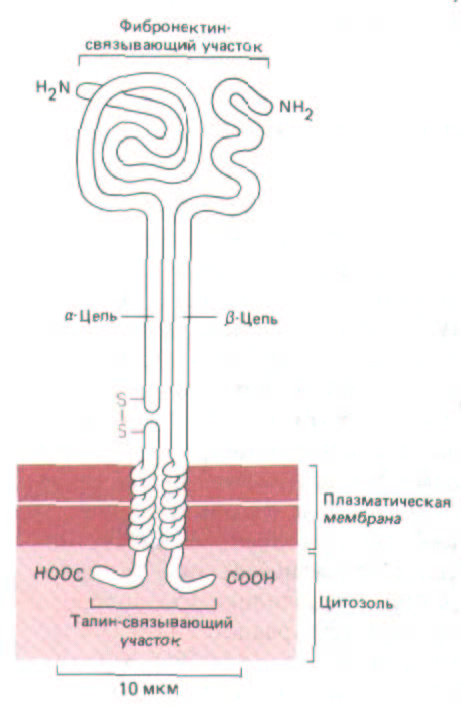
Рис. 14-52. Рецептор фибронектина на клеточной поверхности и его субъединицы. Судя по электронным микрофотографиям изолированных рецепторов, форма молекулы близка к изображенной на рисунке. Глобулярная «головка» выступает над липидным бислоем более чем на 20 нм. Связываясь с фибронектином снаружи и с цитоскелетом внутри клетки (через прикрепительный белок талин), рецептор действует как трансмембранный линкер. Обе цепи, α и β, гликозилированы (на схеме не показано) и удерживаются вместе нековалентными связями.
Первоначально α-цепь синтезируется в виде одной полипептидной цепи с мол. весом 140000, которая затем расщепляется на малую трансмембранную цепь и большую внеклеточную цепь, соединенные дисульфидной связью. Внеклеточная часть β-цепи состоит из повторяющихся участков, богатых цистеином, что указывает на большое число межцепочечных дисульфидных связей (не показаны). Рецептор фибронектина принадлежит к обширному суперсемейству гомологичных рецепторов для матрикса, называемых интегринами, большинство из которых узнает последовательности RGD связываемых ими внеклеточных белков.
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
cyte function associated), другой -Mac-1, так как он в основном встречается на макрофагах. Эти рецепторы участвуют как в межклеточных взаимодействиях, так и во взаимодействиях клеток с матриксом; они играют решающую роль в способности этих клеток бороться с инфекцией.
Люди с наследственным дефицитом лейкоцитарной адгезии неспособны синтезировать β-субъединицу; поэтому их лейкоциты лишены целого семейства рецепторов, и больные подвержены повторным бактериальным инфекциям. Ряд гликопротеинов клеточной поверхности, участвующих в позиционно-специфической клеточной адгезии у личинок Drosophila, тоже принадлежат к суперсемейству интегринов, однако их отношение к трем семействам, свойственным млекопитающим, неизвестно.
Однако не все рецепторы для матрикса принадлежат к этому суперсемейству. Например, некоторые клетки, по-видимому, используют для прикрепления к коллагену неродственный трансмембранный гликопротеин, а у многих клеток, как уже упоминалось, имеются интегральные мембранные протеогликаны, прикрепляющие клетки к внеклеточному матриксу.
14.2.18. Цитоскелет и внеклеточный матрикс взаимодействуют через плазматическую мембрану [26]
Макромолекулы внеклеточного матрикса оказывают поразительное воздействие на поведение клеток в культуре, влияя не только на их движение, но и на форму, полярность, метаболизм и дифференцировку. Например, клетки эпителия роговицы при росте на искусственных поверхностях производят очень мало коллагена; если же культивировать их на ламинине, коллагене или фибронектине, то они накапливают и секретируют коллаген в больших количествах. Другие примеры влияния внеклеточного матрикса на метаболизм и дифференцировку клеток будут обсуждаться в гл. 17 (разд. 17.7.1).
Матрикс может также влиять на организацию цитоскелета клетки. Обычно базальные поверхности эпителиальных клеток, растущих на пластике или стекле, имеют неправильную форму, а прилегающий к ним изнутри цитоскелет дезорганизован. Но когда те же клетки растут на подложке из подходящих макромолекул внеклеточного матрикса, базальные поверхности становятся гладкими, а цитоскелет над ними - таким же упорядоченным, как в интактной ткани. Сходные результаты были получены на культурах фибробластов, подвергшихся опухолевой трансформации. Трансформированные клетки часто вырабатывают меньше фибронектина, чем нормальные культивируемые клетки, и отличаются от них поведением: например, они слабо прикрепляются к субстрату и неспособны распластываться на нем или формировать организованные внутриклеточные пучки актиновых филаментов, известных под названием стрессовых волокон (разд. 11.1.17). У некоторых из таких клеток недостаток фибронектина по крайней мере частично ответствен за их аномальное поведение: если клетки растут на матриксе из организованных волокон фибронектина, то они распластываются и формируют внутриклеточные стрессовые волокна, лежащие параллельно волокнам внеклеточного фибронектина.
Взаимодействие между внеклеточным матриксом и цитоскелетом бывает двусторонним: внутриклеточные актиновые филаменты могут влиять на расположение секретируемых молекул фибронектина. Например, в культуре поблизости от фибробластов волокна внеклеточного фибронектина выстраиваются по направлению смежных внутриклеточных стрессовых волокон (рис. 14-53). Если такие клетки обработать цитохалазином, который разрушает внутриклеточные актиновые фила-
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
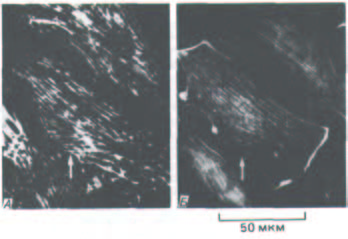
Рис. 14-53. Иммунофлуоресцентные микрофотографии внеклеточных волокон фибронектина (А) и внутриклеточных пучков актиновых филаментов (Б) в трех культивируемых фибробластах крысы. Для выявления фибронектина использованы антитела к фибронектину с присоединенным родамином, а для выявления актина-антитела к актину с присоединенным флуоресцеином. Обратите внимание, что направление волокон фибронектина совпадает с направлением пучков актиновых нитей. (R. О. Hynes, А. Т. Destree. Cell 15: 875-886, 1978. Copyright Cell Press.)
менты, то волокна фибронектина отделяются от клеточной поверхности (точно так же, как во время митоза, когда клетка округляется). Ясно, что должна существовать связь между внеклеточным фибронектином и внутриклеточными актиновыми филаментами через плазматическую мембрану фибробласта. Такую связь осуществляют рецепторы фибронектина, о которых уже говорилось, что эти трансмембранные белки соединяют фибронектин с актиновыми филаментами через такие внутриклеточные прикрепительные белки, как талин (см. разд. 11.2,8. и рис. 14-52). Участок рецептора, связывающий талин, содержит остаток тирозина, фосфорилирование которого тирозин-специфической протеинкиназой, по-видимому, инактивирует этот участок и таким образом разрушает связь между фибронектином и кортикальными актиновыми филаментами. Как полагают, прикрепление клеток к матриксу может регулироваться таким путем специфическими факторами роста, активирующими тирозин-специфические киназы (см. рис. 13-37).
Поскольку цитоскелет клеток способен упорядочивать секретируемые ими макромолекулы матрикса, а те в свою очередь организуют цитоскелет контактирующих с ними клеток, внеклеточный матрикс может в принципе распространять упорядочивание от клетки к клетке (рис. 14-
54). Таким образом, можно полагать, что внеклеточный матрикс играет центральную роль в создании и поддержании ориентации клеток в тканях и органах в процессе развития; например, параллельное расположение фибробластов и коллагеновых волокон в сухожилии мог бы частично отражать именно такие взаимодействия между клетками и матриксом. Трансмембранные рецепторы для матрикса служат в этом процессе упорядочивания «посредниками».
Заключение
Клетки в соединительных тканях погружены в сложный внеклеточный матрикс, который не только скрепляет клетки и ткани, но и
влияет на развитие, полярность и поведение контактирующих с ним клеток. Матрикс содержит различные волокнообразующие белки, вплетенные в гидратированный гель, состоящий из сети цепочек гликозаминогликанов. Гли-

Рис. 14-54. Гипотетическая схема передачи упорядоченности от клетки к клетке через внеклеточный матрикс. Для простоты на рисунке показано, как одна клетка влияет на ориентацию соседних клеток, но та же схема позволяет объяснить и взаимное влияние клеток друг на друга.
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
козамшогликаны представляют собой разнородную группу длинных, отрицательно заряженных полисахаридных цепей, которые (за исключением
гиалуроновой кислоты) ковалентна связаны с белками, образуя молекулы протеогликанов.
Существуют волокнообразующие белки двух функциональных типов: преимущественно структурные (коллаген и эластин) и главным
образом адгезивные (такие, как фибронектин и ламинин). Фибриллярные коллагены (типы I, II и III) представляют собой канатовидные
трехспиральные молекулы, которые во внеклеточном пространстве агрегируют в длинные фибриллы, а те в свою очередь могут
организовываться в разнообразные высокоупорядоченные структуры. Молекулы коллагена типа IV организуются в пластоподобные сети, составляющие основу всех базальных мембран. Молекулы эластина благодаря многочисленным поперечным сшивкам образуют сеть волокон и
слоев, которые могут растягиваться и вновь сокращаться, придавая матриксу упругость. Фибронектин и ламинин служат примерами крупных
адгезивных гликопротеинов матрикса; фибронектин очень широко распространен в соединительных тканях, а ламинин содержится главным
образом в базальной мембране. Благодаря своим множественным прикрепительным доменам такие белки способствуют клеточной адгезии и
участвуют в организующем влиянии внеклеточного матрикса на клетки. Многие из этих адгезивных гликопротеинов содержат общую
трипептидную последовательность (RGD), которая составляет часть структуры, узнаваемой интегринами - членами суперсемейства
гомологичных трансмембранных рецепторов для компонентов матрикса.
Все белки и полисахариды матрикса локально секретируются клетками, соприкасающимися с матриксом; в тесном взаимодействии с
наружной поверхностью плазматической мембраны эти молекулы могут упорядочиваться. Поскольку структура и ориентация матрикса в свою
очередь влияет на ориентацию контактирующих с ним клеток, весьма вероятно, что упорядоченность будет распространяться по матриксу от
клетки к клетке.
14.3. Межклеточное узнавание и адгезия [27]
До сих пор мы рассматривали, как межклеточные соединения и внеклеточный матрикс удерживают клетки вместе в зрелых тканях и органах. Но каким образом клетки объединяются друг с другом на начальных стадиях формирования тканей? Существуют по меньшей мере два принципиально различных способа. Чаще всего ткань образуется из «клеток-основательниц», потомки которых остаются вместе просто потому, что они прикреплены к макромолекулам внеклеточного матрикса и/или к другим клеткам (рис. 14-55). Конкретные особенности таких соединений и определяют структуру клеточного ансамбля. Эпителиальные клеточные пласты обычно возникают именно таким путем, и процессы эмбрионального развития животных в значительной части сводятся к формированию, изгибанию и дифференцировке таких клеточных пластов, что приводит к созданию тканей и органов взрослого организма. Как правило, все клетки раннего зародыша организованы в эпителии, и только позже некоторые клетки изменяют свои адгезивные свойства, выходят из пластов и формируют ткани других типов (разд. 16.1.4-16.1.II).
Другая стратегия формирования ткани представляется более сложной и включает миграцию клеток: одна клеточная популяция проникает в другую и объединяется с ней (а иногда и с другими мигрирующими клетками), формируя ткань смешанного происхождения. Например, в зародышах позвоночных клетки нервного гребня выселяются из эпи-
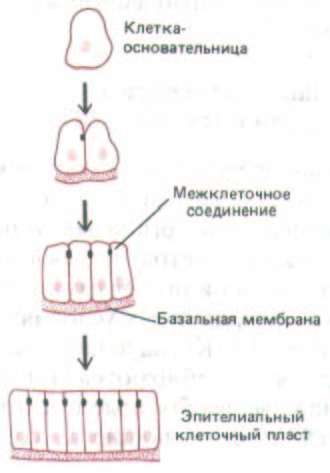
Рис. 14-55. Простейший механизм образования ткани из клеток. Потомки клеток-основательниц удерживаются в эпителиальном слое с помощью базальной мембраны и механизмов межклеточной адгезии (включая специализированные межклеточные соединения).
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
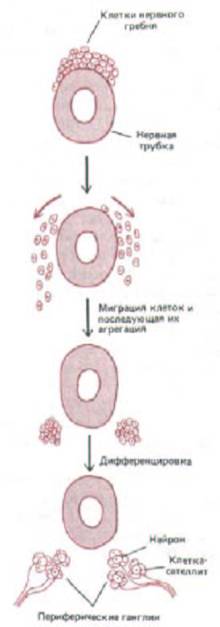
Рис. 14-56. Пример более сложного механизма построения ткани из клеток. Клетки нервного гребня мигрируют из эпителия на верхней поверхности нервной трубки и направляются во многие другие участки зародыша, где образуют различные группы клеток и ткани. Здесь показано, как эти клетки объединяются и дифференцируются в два скопления нейронов периферической нервной системы. Такие скопления называют ганглиями. Другие клетки нервного гребня в ганглии дифференцируются в опорные (сателлитные) клетки, окружающие нейроны.
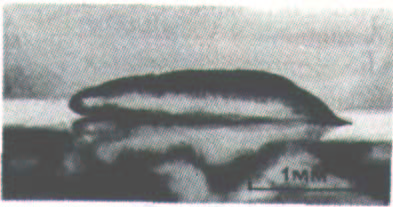
Рис. 14-57, Световая микрофотография ползущего плазмодия миксомицета Dictyostelium discoideum. (С любезного разрешения David Francis.)
телиальной (нервной) трубки, в состав которой они первоначально входили, и по определенным путям мигрируют во многие другие участки. Там они группируются и дифференцируются в различные ткани, в том числе и элементы периферической нервной системы (рис. 14-56). Для такого процесса нужен какой-то механизм, направляющий клетки к месту их назначения, например секреция растворимого химического агента, привлекающего мигрирующие клетки (путем хемотаксиса), или отложение во внеклеточном матриксе адгезивных молекул типа фибронектина (разд. 14.2.13), направляющих миграцию клеток по определенным путям (путем контактной ориентировки).
Достигнув места назначения, мигрирующая клетка должна узнавать другие клетки соответствующего типа, чтобы формировать вместе с ними ткань. Даже в тканях, образующихся без миграции, составляющие их клетки, по-видимому, специфически узнают друг друга: если такую развивающуюся ткань диссоциировать на отдельные клетки, то они предпочтительно вновь ассоциируют друг с другом, а не с клетками другой ткани (разд. 14.3,4). По-видимому, такое специфическое межклеточное узнавание способствует тому, что клетки развивающейся ткани остаются в контакте друг с другом и отделены от клеток соседних тканей.
В попытках понять, как узнают друг друга клетки в развивающихся животных тканях, были проведены остроумные эксперименты на некоторых простых микроорганизмах, способных переходить от одноклеточного существования к многоклеточному и обратно. Независимо от их значения как возможных моделей межклеточного взаимодействия у животных эти организмы интересны и сами по себе.
14.3.1. Миксамебы слизевика при голодании агрегируют с образованием многоклеточных плодовых тел [28]
Слизевик (миксомицет) Dictyostelium discoideum представляет собой эукариотический организм, геном которого только в 10 раз больше, чем у бактерии, и в 100 раз меньше, чем у человека. Эти организмы живут в лесной подстилке в виде отдельных подвижных клеток, называемых миксамебами, питающихся бактериями и дрожжами и при оптимальных условиях делящихся раз в несколько часов (в лабораторных условиях их можно выращивать в жидкой синтетической среде). Когда запасы пищи истощаются, миксамебы перестают делиться и собираются вместе, образуя крошечные (1-2 мм) многоклеточные червеобразные существа (плазмодии), ползающие наподобие слизней и оставляющие за собой след из слизи (рис. 14-57).
Каждый плазмодий формируется путем агрегации до 100000 клеток и проявляет черты поведения, не свойственные свободноживущим миксамебам. Например, плазмодий чрезвычайно чувствителен к свету и теплу и может мигрировать по направлению к такому слабому источнику света, как флуоресцирующий циферблат часов; по-видимому, такое поведение помогает ему двигаться в направлении более благоприятных условий. По мере движения клетки приступают к дифференцировке,
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
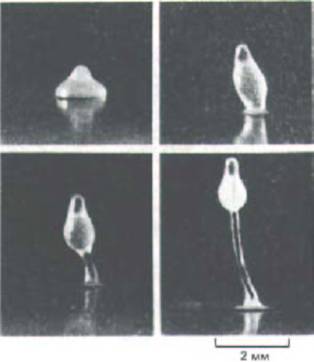
Рис. 14-58. Разные стадии образования плодового тела Dictyostelium discoideum. (Световые микрофотографии; с любезного разрешения John Bonner.)
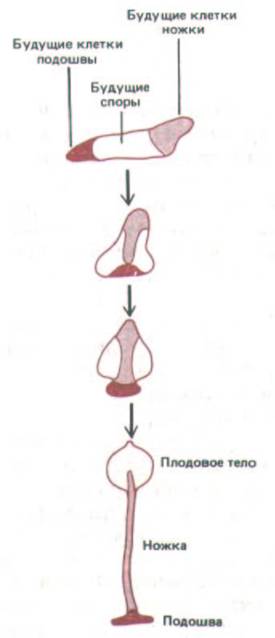
Рис. 14-59. Миграции клеток при формировании плодового тела у Dictyostelium discoideum. Клетки передней части слизевика перемещаются вниз и образуют ножку, а клетки средней части мигрируют вверх и дифференцируются в споры, образующие плодовое тело.
результатом которой примерно через 30 ч после начала агрегации будет образование миниатюрной, напоминающей растение структуры, состоящей из ножки и плодового тела (рис. 14-58). В плодовом теле содержится множество спор, которые могут долгое время выживать даже в крайне неблагоприятных условиях. На рис. 14-59 схематически показаны сложные миграции клеток, происходящие при формировании ножки и плодового тела. Клетки на переднем конце плазмодия становятся участком ножки, следующие за ними дифференцируются в споры, а замыкающие-в подошву.
И клетки ножки, и споры покрываются внеклеточным матриксом (в виде целлюлозных стенок), и в конце концов все клетки, за исключением спор, погибают. Только при наступлении благоприятных условий споры прорастают в свободноживущих миксамеб, возобновляющих цикл (рис. 14-60).
14.3.2. Амебы слизевика агрегируют в результате хемотаксиса [29]
При формировании плазмодия отдельные клетки слизевика агрегируют в результате хемотаксиса, который нам придется рассмотреть, прежде чем обсуждать роль межклеточной адгезии. Одна из реакций на голодание у миксамеб состоит в том, что они начинают вырабатывать и выделять сАМР, который служит хемотаксическим сигналом, привлекающим других миксамеб. (Как мы знаем из гл. 12, в прокариотических и животных клетках сАМР служит внутриклеточным сигналом; Dictyostelium-единственный организм, у которого он действует еще и как внеклеточная сигнальная молекула.) По-видимому, агрегация инициируется случайным образом: любые клетки, начинающие первыми секретировать циклический AMP, привлекают другие клетки и таким образом становятся центрами агрегации. Циклический AMP, вырабатываемый такими «клетками-инициаторами», секретируется отдельными «импульсами» и связывается специфическими рецепторами на поверхности соседних голодающих амеб, направляя тем самым их движение
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
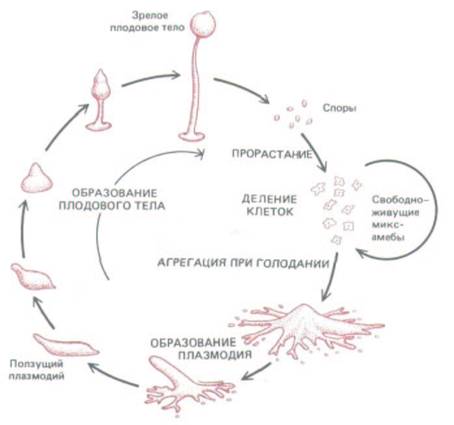
Рис. 14-60. Жизненный цикл Dictyostelium discoideum. При голодании свободноживущие миксамебы агрегируют с образованием подвижного плазмодия, который затем образует плодовое тело. При благоприятных условиях высвобождающиеся из плодового тела споры прорастают и вновь превращаются в амеб.
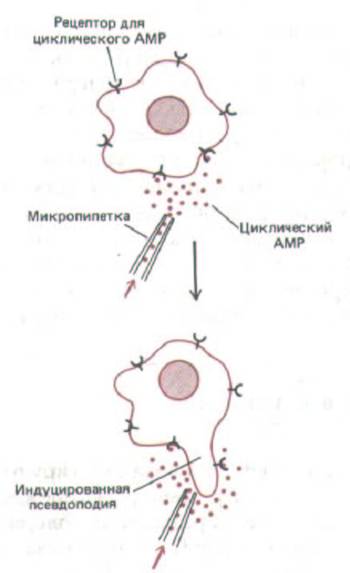
Рис. 14-61. Нанесение небольшого количества циклического AMP на любую точку поверхности голодающей клетки (амебы) Dictyostelium
тотчас же вызывает образование псевдоподии в этой точке. Такой механизм позволяет амебе двигаться по направлению к источнику сАМР. Чтобы воздействовать на клетку, циклический AMP должен связаться со специфическими рецепторами на ее поверхности.
в сторону источника циклического AMP. Такой хемотаксический ответ можно продемонстрировать, нанеся из микропипетки ничтожное количество сАМР на любой участок поверхности клетки голодающей миксамебы. Ответом будет немедленное образование псевдоподии, растущей в сторону пипетки (рис. 14-61); псевдоподия прикрепляется к поверхности, на которой находится клетка, и тянет клетку в том же направлении. Как только образуется центр агрегации, зона его влияния быстро расширяется, так как агрегирующие клетки не только отвечают на сигнал циклического AMP, но и передают его от клетки к клетке. Каждый импульс циклического AMP побуждает соседние клетки не только к движению к источнику импульса, но и к испусканию собственного импульса в виде циклического AMP. Этот новый, высвобождающийся с небольшой задержкой импульс в свою очередь ориентирует находящиеся рядом клетки, вызывая у них тоже выброс сАМР, и т.д. Таким образом возникают регулярные чередующиеся волны циклического AMP, распространяющиеся из каждого центра агрегации, заставляя более удаленных миксамеб двигаться внутрь концентрическими или спиральными волнами, которые можно видеть на кадрах цейтраферных фильмов (рис. 14-62). Преимущество такой системы передачи состоит в том, что по мере распространения из центра сигнал постоянно возобновляется, не ослабляясь на большом расстоянии.
В отличие от этого сигнал, распространяющийся только путем диффузии, постепенно ослабевает по мере распространения. Это различие можно ясно увидеть, сравнивая процесс агрегации у Dictyostelium discoideum и у D. minutum- формы, у которой релейная система передачи отсутствует. У D. munutum уровень сигнала, исходящего из каждого центра агрегации, сильно ослабевает, и в результате формируются очень мелкие плазмодии и плодовые тела.
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
14.3.3. Межклеточная адгезия у слизевиков зависит от специфических гликопротеинов клеточной поверхности [30]
Помимо активации сигнальной системы циклического AMP голодание миксамеб Dictyosteliun вызывает экспрессию сотен новых генов, и некоторые из них кодируют молекулы межклеточной адгезии, участвующие в агрегации клеток. Полагают, например, что один из связывающих углеводы белков (т.е. лектинов, см. разд. 6.3.1)- дискоидин-1 -выделяет-ся голодающими клетками для обеспечения примитивных форм контактной ориентировки. Связываясь с поверхностью миксамебы и с субстратом, по которому она мигрирует, он мог бы способствовать образованию потоков миксамеб, движущихся к центрам агрегации, во многом подобно тому, как фибронектин направляет миграцию клеток во время гаструляции. В самом деле, связывание клетки с дискоидином-1 зависит от того же трипептида RGD, который содержится в фибронектине и многих других адгезивных белках (разд. 14.2.13).
Различные вновь синтезируемые белки способствуют процессу межклеточной адгезии, позволяя мигрирующим миксамебам плотно слипаться друг с другом и формировать многоклеточный организм. В первые 8 ч голодания клетки слипаются с помощью Са2 +-зависимого механизма с участием адгезивной молекулы, называемой контактным сайтом В. Через 8 ч вступает в действие другая адгезионная система, где слипание клеток осуществляется Са2 +-независимым механизмом с участием молекулы межклеточной адгезии, называемой контактным сайтом А.
Контактные сайты А и В были выделены и идентифицированы как интегральные гликопротеины плазматической мембраны с помощью остроумного иммунологического метода, представленного на рис. 14-63. Позднее этот метод был использован для идентификации молекул межклеточной адгезии также и у позвоночных.
Каким образом гликопротеины клеточной поверхности, такие как контактные сайты А и В, связывают клетки друг с другом? На рис. 14-
64 представлены три возможных механизма: 1) молекулы одной клетки могут связываться с такими же молекулами соседних клеток (так называемое гомофильное связывание); 2) молекулы одной клетки могут связываться с иного рода молекулами соседних клеток (гетерофильное
связывание); и 3) рецепторы клеточной поверхности соседних клеток могут связываться друг с другом, секретируя мультивалентные линкерные молекулы. Как выяснилось, у животных действуют все эти три механизма.
Полагают, что контактный сайт А обеспечивает слипание клеток путем гомофильного механизма, так как после присоединения белка к синтетическим агрегатам эти агрегаты связываются только с клетками, вырабатывающими контактный сайт А, и это связывание блокируется, если клетки предварительно обработаны антителами к контактному сайту А. Секвенирование ДНК показывает, что контактный сайт А - это лишь один раз пронизывающий мембрану белок, по-видимому, не родственный ни одному из до сих пор известных белков межклеточной адгезии (см. ниже).
|
|
|
|
|
Дата добавления: 2015-08-31; Просмотров: 2910; Нарушение авторских прав?; Мы поможем в написании вашей работы!