КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Обработка зрительной информации
|
|
|
|
Фоторецепция
Физические основы зрения человека. Зрение представляет собой восприятие электромагнитных излучений сравнительно узкого диапазона, то есть с длиной волны от 400 нм до 750 нм (при длине волны, равной 435 нм, возникает ощущение синего цвета, 546 нм - зеленого, 589 нм - желтого, 671 нм - красного). Для восприятия разной длины света в сетчатки имеются соответствующие зрительные пигменты, находящиеся в палочках или колбочках. Пигмент родопсин (преимущественно содержится в палочках) имеет максимум поглощения для излучения с длиной волны в 500 нм, йодопсин, хлоролаб и эритролаб (пигменты колбочек) - для излучений с длиной волной соответственно 560 нм, 546 нм и 671 нм. При этом орган зрения приспособлен к восприятию различного по интенсивности светового потока. Если яркость среды выразить в канделах (единица измерения интенсивности света) на м2, то ночью без луны и звезд яркость среды составляет 10-6 кд/м2, ночью со звездами -10-3 кд/м2, ночью при полной луне -10-1кд/м2, а в яркий солнечный день - 107 кд/м2. Таким образом, диапазон изменения интенсивности освещенности, воспринимаемой глазом, огромен (от 10-6 до 107 кд/м2).
Пигментный слой сетчатки. Он образован одним рядом эпителиальных клеток (пигментных эпителиоцитов), лежащих на базальной мембране. От внутренней поверхности пигментных клеток отходит по 8-10 цитоплазматических отростков («борода»), заполненных пигментом меланином, или фусцином (он синтезируется меланосомами эпителиоцитов), отделяющих друг от друга наружные сегменты палочек и колбочек. Благодаря меланину доходящий до сетчатки свет не рассеивается и не отражается, а полностью попадает на фоторецепторы, что способствует четкости зрительного восприятия. Эти же клетки поглощают избыточное количество квантов света при наличии чрезмерной освещенности сетчатки (при сильном освещении зерна пигмента перемещаются из эпителиальных клеток и заслоняют палочки и колбочки от яркого света). Кроме того, эпителиальные клетки пигментного слоя принимают участие в ресинтезе зрительного пигмента фоторецепторов после его обесцвечивания, а также (за счет фагоцитоза) в удалении отработанных наружных сегментов зрительных клеток. С помощью пигментных эпителиоцитов происходит перенос к фоторецепторам кислорода и питательных веществ. Нарушение всех этих функций пигментного слоя, которое наблюдается при отслойке сетчатки, вызывает дегенерацию фоторецепторов, что приводит к развитию слепоты.
У ночных животных между пигментными клетками и фоторецепторами расположен слой, отражающий свет. Он состоит из особых кристаллов или нитей. В результате отражения света от кристаллов у ночных животных при внешнем освещении светятся глаза. В этом случае на фоторецепторы действуют не только прямые лучи, но и отраженные, что в условиях слабой освещенности повышает возможность восприятия света.
Восприятие света осуществляется с участием фоторецепторов, или нейросенсорных клеток, которые относятся ко вторичночувствующим рецепторам. Это означает, что они представляют собой специализированные клетки, передающие информацию о квантах света на нейроны сетчатки, в том числе вначале на биполярные нейроны, затем на ганглиозные клетки, аксоны которых составляют волокна зрительного нерва; информация затем поступает на нейроны подкорковых (таламус и передние бугры четверохолмия) и корковых центров (первичное проекционное поле 17, вторичные проекционные поля 18 и 19) зрения. Кроме того, в процессах передачи и переработки информации в сетчатке участвуют также горизонтальные и амакриновые клетки. Все нейроны сетчатки образуют нервный аппарат глаза, который не только передает информацию в зрительные центры мозга, но и участвует в ее анализе и переработке. Поэтому сетчатку называют частью мозга, вынесенной на периферию.
Более 100 лет назад на основании морфологических признаков Макс Шультце разделил фоторецепторы на два типа - палочки (длинные тонкие клетки, имеющие цилиндрический наружный сегмент и равный ему по диаметру внутренний) и колбочки (обладающие более коротким и толстым внутренним сегментом). Он обратил внимание на то, что у ночных животных (летучая мышь, сова, крот, кошка, еж) в сетчатке преобладали палочки, а у дневных (голуби, куры, ящерицы) - колбочки. На основании этих данных Шультце предложил теорию двойственности зрения, согласно которой палочки обеспечивают скотопическое зрение, или зрение при низком уровне освещенности, а колбочки реализуют фотопическое зрение и работают при более ярком освещении. Следует, однако, отметить, что кошки прекрасно видят днем, а содержащиеся в неволе ежи легко приспосабливаются к дневному образу жизни; змеи, в сетчатке которых находятся главным образом колбочки, хорошо ориентируются в сумерках.
В настоящее время показано, что ночное, или палочковое, зрение имеет высокую чувствительность (в 1000 раз выше, чем колбочковое), низкую разрешающую способность (остроту) и является черно-белым (ахроматическим). Абсолютный порог зрения соответствует одновременному поглощению 6-7 фотонов сетчаткой, содержащей около 50 палочек. Палочковое зрение не цветное, так как все палочки содержат один зрительный пигмент - родопсин, и, следовательно, различия в длине волны воспринимаются как различия в интенсивности. Палочковое зрение неотчетливо потому, что конвергенция большого числа палочковых клеток на одно нервное волокно делает эффективную мозаику сетчатки довольно грубой. В целом, сегодня не вызывает сомнения, что палочковые фоторецепторы, расположенные в основном по периферии сетчатки (поэтому в сумерки лучше видны предметы, расположенные по сторонам) обеспечивают скотопическое и мезопическое зрение, то есть зрение при слабом дневном свете и при сумерках соответственно, и они предназначены для детектирования интенсивности света. Нарушение функции палочек вызывает расстройство сумеречного зрения - так называемую куриную слепоту: человек совершенно слепнет в сумерках, но днем зрение остается нормальным (обычно возникает при недостатке в пище витамина А).
Колбочковые фоторецепторы (три варианта) преимущественно сконцентрированы в области центральной ямки сетчатки (до 140 тыс. колбочек на 1 мм2). Они обеспечивают фотопическое зрение, т.е. зрение при обычном дневном свете, при котором человек способен различать цвета. Дневное, или колбочковое, зрение обладает более низкой абсолютной чувствительностью, чем палочковое в силу того, что колбочки по своей природе мене чувствительны и, кроме того, к одной ганглиозной клетке их сходится меньше, чем палочек. Однако уменьшение конвергенции приводит к повышению остроты зрения. Колбочковое зрение может быть цветным при наличии нескольких типов колбочек с различными зрительными пигментами, поглощающими в различных областях спектра. Поражение колбочек вызывает светобоязнь: человек видит при слабом свете, но слепнет при ярком освещении. В этом случае может развиться и полная цветовая слепота - ахромазия. Кроме того, отсутствие одного из видов колбочек приводит к нарушению цветоощущения, т.е. к дальтонизму.
Морфологические особенности палочек и колбочек. В сетчатке человека в каждом глазу содержится около 110-123 млн. палочек и примерно 6-7 млн. колбочек, т.е. всего 130 млн. фоторецепторов. В области желтого пятна имеются главным образом колбочки, а на периферии - палочки.
Все фоторецепторы (и палочки, и колбочки) состоят из двух сегментов (члеников) - наружного, светочувствительного, и внутреннего, обеспечивающего метаболизм фоторецепторов. Между собой наружный и внутренний сегменты связаны соединительной ножкой (это пучок из 16-18 тонких фибрилл).
Наружные (светочувствительные) сегменты фоторецепторов обращены в сторону, противоположенную свету (наибольшее возбуждение от действия света наблюдается в тех случаях, когда направление луча совпадает с длинной осью палочки или колбочки). У палочек они представляет собой стопку тонких фоторецепторных дисков (примерно от 4000 до 1000 штук) диаметром 6 мкм каждый. Каждый диск - это двойная мембрана, состоящая из двух мономолекулярных слоев липидов, помещающихся между двумя слоями молекул белка. С этими молекулами белка связан зрительный пигмент. У палочек наружный сегмент намного длиннее и более богат зрительным пигментом, чем у колбочек. Это частично объясняет более высокую чувствительность палочки к свету: палочку может возбудить всего один квант света, а колбочку - только около 100 квантов. Фоторецепторные диски постепенно изнашиваются (особенно интенсивно - в палочках), поэтому верхушка наружного сегмента периодически обламывается и фагоцитируется клетками пигментного слоя (в палочках это совершается в утренние и дневные часы, в колбочках - в вечерние и ночные). На замену старым дискам из внутреннего сегмента фоторецептора постоянно поступают новые диски
Внутренний сегмент фоторецепторов содержит множество митохондрий, а также ядро, рибосомы, элементы эндоплазматической сети и пластинчатого комплекса Гольджи. Он является основным местом образования энергии и белков. Именно во внутреннем сегменте непрерывно совершается синтез зрительного пигмента и его упаковка в фоторецепторные диски, которые затем поступают в наружный сегмент фоторецептора для замены устаревших дисков. За 1 час образуется 3 таких диска, перемещение которых в наружный сегмент осуществляется в течение 2-3 недель.
Внутренний сегмент фоторецепторов оканчивается лентовидным образованием. Это пресинаптическое окончание, в котором содержатся везикулы, наполненные глутаматом. Благодаря этому образованию сигнал с фоторецептора передается на контактирующую с ним биполярную клетку.
Принцип фоторецепции. В условиях темноты фоторецепторная клетка (палочковая или колбочковая) постоянно выделяет медиатор глутамат, который гиперполяризует биполярную клетку, соединенную с ганглиозной клеткой. Поэтому в темноте биполярная клетка не возбуждает ганглиозную клетку и потенциалы действия не идут по зрительному нерву к подкорковым и корковым зрительным центрам. При попадании кванта света происходит «возбуждение» фоторецепторной клетки. Оно проявляется в том, что внутренний сегмент фоторецепторной клетки гиперполяризуется и тем самым снижает выделение медиатора (глутамата), т.е. происходит снятие тормозного влияния глутамата на биполярную клетку. Это приводит к тому, что биполярная клетка возбуждается и в свою очередь (за счет выделения возбуждающего медиатора, в роли которого выступает ацетилхолин) активирует ганглиозную клетку, благодаря чему генерируются потенциалы действия, которые по зрительному нерву достигают подкорковых и корковых зрительных центров, что в конечном итоге создает возможность для восприятия зрительного образа. Наличие в сетчатке горизонтальных (они регулируют синаптическую передачу между фоторецепторами и биполярными клетками) и амакриновых клеток (регулирующих взаимодействие между биполярными клетками и ганглиозными и осуществляющими процессы латерального торможения) позволяет создать оптимальные условия для детекции светового сигнала. Не исключено, что взаимодействие между нейронами сетчатки регулируется и со стороны головного мозга, так как в составе зрительного нерва обнаруживаются эфферентные волокна.
Современные представления о процессах, происходящих в фоторецепторной клетке при воздействии света. В наружных сегментах палочек сетчатки человека содержится зрительный пигмент родопсин, или зрительный пурпур, максимум спектра поглощения которого находится в области 500 нм. Его молекулярная масса составляет 40 кД. Он относится к каротиноидам, т.е. к хромолипопротеинам. Поглощение света таким белком обеспечивается наличием хромофорной группы атомов, в качестве которой выступают альдегиды спиртов витамина А, их называют ретиналями. Во всех зрительных пигментах ретиналь всегда находится в 11-цисформе, у которой длинная часть цепочки изогнута и скручена в весьма нестабильную конформацию (такая форма называется 11- цис-ретиналем). В норме 11-цис-ретиналь связан с бесцветным белком опсином. Это соединение и является родопсином, который в той или иной модификации служит универсальным молекулярным медиатором фоторецепции у всех животных. Источником ретиналя в организме являются каротиноиды. Поэтому их недостаток приводит к дефициту витамина А, а как следствие, к недостаточному ре-синтезу родопсина, что в свою очередь является причиной нарушения сумеречного зрения («куриной слепоты»).
При поглощении кванта света родопсином происходит мгновенная цистрансизомеризация ретиналя, т.е. 11-цис-ретиналь выпрямляется и превращается в полностыо-транс-ретиналь. В результате этой реакции изомеризации, которая длится около 1 пс (10-12 с), происходит обесцвечивание родопсина, т.е. образование метародопсина II (последовательность событий: родопсин → прелюмиродопсин → люмиродопсин → метародопсин I → метародопсин II; при этом метародопсин I имеет темно-оранжевый цвет, а метародопсин II - светло-желтый.)
В свою очередь образовавшийся метародопсин II взаимодействует с трансдуцином (в соотношении 1 молекула метародопсина II на 500-1000 молекул трансдуцина). Трансдуцин представляют собой разновидность G-белка, т.е. гуанозинтрифосфат-связывающего белка, который широко представлен в различных клетках как посредник в передаче сигнала от мембранного рецептора к внутриклеточному эффектору (например, от β-адренорецепторов к кальциевому насосу). В комплексе с метародопсином II трансдуцин переходит в активное состояние и обменивает связанный с ним в темноте гуанозиндифосфат (ГДФ) на гуанозинтрифосфат (ГТФ). В свою очередь активированный трансдуцин, связанный с молекулой ГТФ, активирует фермент фосфодиэстеразу (ФДЭ) - в соотношении 1 молекула трансдуцина - 1 молекула ФДЭ. Активированная ФДЭ с высокой скоростью разрушает молекулы цГМФ, т.е. циклического гуанозинмонофосфата (1 молекула ФДЭ разрушает несколько тысяч молекул цГМФ), что приводит к существенному снижению концентрации свободного цГМФ в цитоплазме наружного сегмента рецептора. Так как именно цГМФ держит натриевые и кальциевые каналы наружного сегмента фоторецептора в открытом состоянии (это происходит в темноте), то снижение концентрации свободной цГМФ приводит к закрытию натриевых и кальциевых каналов, что в свою очередь порождает гиперполяризацию (т.е. генерацию рецепторного потенциала). Эта гиперполяризация тормозит выделение в пресинаптическом окончании фоторецепторной клетки глутамата, который в условиях темноты вызывал гиперполяризацию биполярной клетки. Тем самым происходит возбуждение биполярной клетки, которое, в свою очередь, передается на ганглиозную клетку (за счет выделения возбуждающего медиатора из пресинаптического окончания биполярной клетки, функцию которого, вероятнее всего, выполняет ацетилхолин). В ответ на генераторный потенциал ганглиозная клетка генерирует потенциалы действия, бегущие по зрительному нерву в подкорковые и корковые зрительные центры.
В связи с тем, что в наружном сегменте фоторецептора при действии света снижается содержание свободного кальция (из-за закрытия кальциевых каналов), это вызывает активацию фермента гуанилатциклазы, благодаря чему усиливается образование цГМФ из ГТФ. Повышение внутриклеточного содержания цГМФ вызывает открытие натриевых и кальциевых каналов в наружном сегменте фоторецептора, что приводит к деполяризации и восстановлению продукции глутамата. Одновременно накопление
цГМФ нарушает взаимодействие метародопсина II с трансдуцином. Таким образом, за короткий промежуток времени фоторецептор восстанавливает свою способность повторно отвечать на световой сигнал.
Отщепившийся от трансдуцина метародпосин II распадается на 11-транс-ретиналь и опсин. В последующем (в условиях темноты) происходит восстановление родопсина, т.е. соединение белка опсина с 11-цис-ретиналем. Ресинтез родопсина осуществляется с участием пигментного эпителия сетчатки при наличии ретинола, т.е. витамина А, а также фермента редуктазы. Редуктаза, с одной стороны, способствует превращению 11-транс-ретиналя в ретинол, т.е. в витамин А, а с другой - катализирует реакцию изомеризации, т.е. образование из витамина А его альдегида (11- цис-ретиналя), который в условиях темноты соединяется с опсином. Так завершается процесс ресинтеза родопсина.
В наружных сегментах трех типов колбочек (сине-, зелено- и красночувствительных) содержится три типа зрительных пигментов, максимумы спектров поглощения которых находятся в синей (435 нм), зеленой (546 нм) и красной (671 нм) частях спектра. Красный колбочковый пигмент получил название «йодопсин». Структура йодопсина близка к родопсину. Но в йодопсине 11-цис-ретиналь соединен с другим белком, который отличается от опсина палочек. Степень поглощения света родопсином и йодопсином различна. Родопсин максимально поглощает лучи в сине-зеленой части спектра. Эти лучи в темноте кажутся наиболее яркими. Йодопсин в наибольшей степени поглощает желтый свет.
Процессы анализа и синтеза в нейронах сетчатки. На 130 млн. фоторецепторных клеток (123 млн. палочек и 7 млн. колбочек) приходится только 1 млн. 250 тыс. ганглиозных клеток, аксоны которых образуют зрительный нерв. Это значит, что сигналы от многих фоторецепторов конвергируют через биполярные нейроны к одной ганглиозной клетке. Ганглиозная клетка сетчатки - это первый нейрон «классического» типа в цепи фоторецептор - мозг.
Фоторецепторы, соединенные с одной ганглиозной клеткой, образуют рецептивное поле этой клетки. Рецептивные поля различных ганглиозных клеток частично перекрывают друг друга. Таким образом, каждая ганглиозная клетка суммирует возбуждение, возникающее в большом числе фоторецепторов. Это повышает световую чувствительность, но ухудшает пространственное разрешение. Однако в районе центральной ямки сетчатки каждая колбочка соединена с одной так называемой карликовой биполярной клеткой, с которой соединена также всего одна ганглиозная клетка. Это обеспечивает высокое пространственное разрешение, но резко уменьшает световую чувствительность.
Для возникновения зрительного ощущения необходимо, чтобы световой раздражитель имел некоторую минимальную (пороговую) энергию. Минимальное число квантов света, необходимое для возникновения ощущения света, в условиях темновой адаптации колеблется от 8 до 47. Рассчитано, что одна палочка может быть возбуждена всего 1 квантом света. Таким образом, чувствительность рецепторов сетчатки в наиболее благоприятных условиях световосприятия физически предельна. Одиночные палочки и колбочки сетчатки различаются по световой чувствительности незначительно, однако число фоторецепторов, посылающих сигналы на одну ганглиозную клетку, в центре и на периферии сетчатки различно. Число колбочек в рецептивном поле в центре сетчатки примерно в 100 раз меньше числа палочек в рецептивном поле на периферии сетчатки. Соответственно и чувствительность палочковой системы в 100 раз выше, чем колбочковой.
Все рецептивные поля сетчатки человека имеют концентрический вид (окружности разного диаметра, но в среднем диаметр рецептивного поля ганглиозной клетки, связанной с палочками, составляет 1 мм). Благодаря этому рецептивные поля ганглиозных клеток сетчатки производят поточечное описание изображения на сетчатке: оно отображается очень тонкой мозаикой, состоящей из возбужденных нейронов. В области центральной ямки сетчатки диаметр рецептивного поля наименьший, а на периферии - намного больше, поэтому лишь в центральной ямке имеет место зона наилучшего видения (максимальная острота зрения).
Рецептивные поля построены таким образом (с участием, конечно, тормозных нейронов), что они позволяют, во-первых, оценить - в какой части рецептивного поля находится световой луч, а во-вторых, - что происходит с сетчаткой: освещается она или нет. При этом часть рецептивных полей сетчатки устроена таким образом, что их ганглиозная клетка (on-нейрон) возбуждается в том случае, если освещается круглый центр этого поля, а кольцевая периферия не освещается. Если сразу осветить и центр, и периферию поля, то нейрон не возбуждается. В других рецептивных полях ганглиозная клетка (off-нейрон) будет возбуждаться только в том случае, если освещается периферия. Среди on- и off-нейронов имеются варианты: одни из них (Х-нейроны) реагируют (например, на постоянное освещение центра) непрерывным возбуждением, т.е. непрерывно генерируют ПД - это медленно адаптирующие нейроны, или «тонические» нейроны. Другие (У-нейроны) возбуждаются только в момент включения или выключения светового сигнала - это быстроадаптирующие нейроны, или «фазические» нейроны. Третие (W-нейроны) возбуждаются только в случае, если стимул «движется», т.е. перемещается по сетчатке.
Для ганглиозных клеток сетчатки характерно явление пространственной суммации (при увеличении размера светового пятнышка, вспыхивающего в центре рецептивного поля, ответ ганглиозной клетки увеличивается), а также явление латерального, или бокового торможения, обеспечивающего четкое видение всех мелких деталей предмета (одновременное возбуждение близко расположенных ганглиозных клеток приводит к их взаимному торможению: ответы каждой клетки делаются меньше, чем при одиночном раздражении).
В целом, благодаря такой сложной организации и специализации нейронов сетчатки уже на этом уровне происходит определение таких качеств светового стимула как освещенность, цвет, форма, движение сигнала.
Важную роль в процессах анализа и синтеза в нейронах сетчатки играет так называемая интегративная система нейронов, включающая в себя горизонтальные и амакриновые клетки, которые связывают элементы сетчатки по горизонтали и осуществляют латеральные взаимодействия. Именно благодаря этой системе на одну ганглиозную клетку конвергируют до сотни биполярных клеток и десятки тысяч рецепторов, а один рецептор может быть связан с десятками ганглиозных клеток.
Показано, что горизонтальные, или звездчатые, клетки, которые регулируют связь фоторецепторов с биполярными клетками, не генерируют нервных импульсов, но их мембрана обладает нелинейными свойствами, обеспечивающими безимпульсное проведение сигнала без затухания. Различают два типа горизонтальных клеток в сетчатке: L-тип и С-тип. Клетки L-типа, или яркостные, всегда отвечают гиперполяризацией вне зависимости от длины волны света. Клетки С-типа, или хроматические, в свою очередь бывают двухфазные и трехфазные. Речь идет о том, что ответ клеток С-типа (гиперполяризация или деполяризация) на световой сигнал зависит от длины стимулирующего света. Двухфазные клетки бывают либо красно-зеленые (деполяризуются красным светом, гиперполяризуются зеленым), либо зелено-синие (деполяризуются зеленым светом, гиперполяризуются синим). Трехфазные клетки деполяризуются зеленым светом, а синий и красный вызывает гиперполяризацию мембраны.
Амакриновые клетки, участвующие в модуляции передачи сигнала от биполярных клеток к ганглиозным, генерируют градуальные и импульсные потенциалы. Эти клетки отвечают быстротекущей деполяризацией на включение и выключение света.
Процессы обработки зрительной информации в подкорковых зрительных центрах. Зрительные нервы (II пара черепных нервов) от каждого глаза встречаются у основания мозга, где формируется их частичный перекрест, или хиазма. Перекрест обеспечивает каждое полушарие большого мозга информацией от обоих глаз: в затылочную долю правого полушария поступают сигналы от правых половин каждой сетчатки, а в левое полушарие - от левых половин сетчаток.
После зрительного перекреста зрительные нервы называют зрительными трактами. Они проецируются в ряд мозговых структур. Основное число волокон приходит в таламический подкорковый зрительный центр - латеральное, или наружное, коленчатое тело. Отсюда сигналы поступают (в составе зрительной лучистости, или пучка Грациоле) в первичную проекционную область зрительной зоны коры (стриарная кора, или 17-е поле по Бродману), затем - во вторичную проекционную зону (поле 18 и 19, престриарная кора), а затем - в ассоциативные зоны коры.
Часть волокон от ганглиозных клеток сетчатки направляется к крыше среднего мозга и в передние бугры четверохолмия, в претектальную область и подушку в таламусе (из подушки информация передается на область 18-го и 19-го полей коры).
Претектальная область ответственна за регуляцию диаметра зрачка, а передние бугры четверохолмия связаны с глазодвигательными центрами и высшими отделами зрительной системы. Нейроны передних бугров четверохолмия обеспечивают реализацию ориентировочных (сторожевых) зрительных рефлексов, в том числе с участием тектоспинального пути. Из передних бугров четверохолмия нервные импульсы следуют в ядра глазодвигательного нерва (двигательное и добавочное вегетативное), ин-нервирующие мышцы глаза, а также ресничную мышцу и мышцу, суживающую зрачок. Благодаря этому, в ответ на попадание световых волн в глаз зрачок суживается, а глазные яблоки поворачиваются в направлении пучка света. Часть информации от сетчатки по зрительному тракту поступает к супрахиазматическим ядрам гипоталамуса, обеспечивая реализацию циркадианных (околосуточных) биоритмов.
В передаточных станциях (передние бугры четверохолмия, латеральные коленчатые тела) происходит вычленение зрительной информации и выявление новых качеств, недоступных «примитивной» сетчатке. С этой целью за счет явления конвергенции и дивергенции создаются более сложные рецептивные поля, а также появляются более «обученные», более «смышленые» нейроны, которые возбуждаются на особые сигналы, например, на стимул, траектория движения которого имеет волнообразный характер. В этих передаточных станциях имеет место сохранение топического расположения рецепторов на сетчатке, а также наличие колонок - вертикальных скоплений нейронов, предназначенных для расчленения информации, поступающей от данной области сетчатки, на отдельные составляющие. Например, в латеральном коленчатом теле имеются нейроны, реагирующие на свет и темноту, отвечающие за контраст, реагирующие на определенные цвета, а также на определенное направление движения светового стимула (дирекционные нейроны).
Рецептивные поля нейронов латерального коленчатого тела круглые, но меньшего размера, чем в сетчатке. Ответы нейронов, генерируемые в ответ на вспышку света, здесь короче, чем в сетчатке. В латеральном коленчатом теле происходит взаимодействие афферентных сигналов, пришедших из сетчатки, с эфферентными сигналами из зрительной области коры, а также (через ретикулярную формацию) от слуховой и других сенсорных систем. Эти взаимодействия обеспечивают выделение наиболее существенных компонентов сенсорного сигнала и процессы избирательного зрительного внимания.
Обработка информации в корковых зрительных центрах. Основной анализ зрительной информации совершается нейронами коры, среди которых выделяют простые, сложные, сверхсложные и гностические. Все эти нейроны объединены в вертикальные колонки (глазодоминантные, ориентационные).
Прежде всего, потенциалы действия нейронов наружного коленчатого тела поступают в затылочную часть полушарий большого мозга, где расположена первичная проекционная область зрительной зоны коры (стриарная кора, или поле 17). Афферентные волокна из наружного коленчатого тела оканчиваются в слое IV и в глубине слоя III 17-го поля, которое является центральным полем зрительной коры, а 18-е и 19-е поля - периферическими. Ассоциативные волокна из поля 17 направляются в 18-е и 19-е поля. Между сетчаткой и полем 17 существует упорядоченное топографическое соответствие, благодаря чему карта сетчатки, а, следовательно, и поле зрения проецируется на кору. Центральная ямка, где острота зрения максимальна, занимает большую часть коркового представительства. Таким образом, единственную точную ретинооптическую карту содержит только поле 17. Смежным с ним полям 18 и 19 приписывают нетопографические ассоциативные функции.
В первичной проекционной зоне происходит анализ информации, поступающей одновременно из правого и левого глаз. Как и в других зонах коры, в этой области анализ проводится с участием колонок. Имеются глазодоминантные колонки, анализирующие информацию, идущую либо из правого глаза, либо из левого. Эти колонки соседствуют друг с другом, поэтому, вероятно, между ними происходит обмен информацией, и это позволяет видеть двумя глазами один предмет (бинокулярное зрение).
Среди нейронов колонок коры различают «простые» нейроны, задача которых - выявить контраст, наличие движущегося стимула, т. е. точно такие же задачи, как у нейронов сетчатки (но для нейронов коры рецептивные поля имеют более обобщенный характер). Простые нейроны зрительной коры в целом выполняет свои специфические функции, получая сигналы от определенного участка сетчатки и откликаясь максимальной активностью на определенную форму и ориентацию стимула.
Кроме того, колонки зрительной коры содержат «сложные» и «сверхсложные» нейроны, которые возбуждаются при наличии определенных условий, например, при движении светового сигнала слева направо или снизу вверх (дирекциональные детекторы), либо возбуждаются на определенный цвет, часть нейронов лучше всего отвечает на относительную удаленность объекта от глаз. Важно подчеркнуть, что информация о разных признаках зрительных объектов (форма, цвет, движение) обрабатывается параллельно в разных частях зрительной зоны коры большого мозга.
Таким образом, в первичной проекционной зоне происходит более специализированная и сложная, чем в сетчатке и в латеральном коленчатом теле, переработка информации. Нейроны зрительной зоны коры имеют не круглые, а вытянутые (по горизонтали, вертикали или в одном из косых направлений) рецептивные поля небольшого размера. Благодаря этому нейроны коры способны выделять из цельного изображения отдельные фрагменты линий с той или иной ориентацией и расположением (детекторы ориентации) и избирательно на них реагировать.
Вся информация от нейронов первичного (поле 17) и вторичных (поля 18 и 19) проекционных полей коры передается в передние и задние ассоциативные зоны коры (лобные и теменно-височные области коры), где с участием более «обученных» (гностических) нейронов происходит окончательное формирование образа. В частности, в ассоциативных полях содержатся нейроны, обученные узнавать все буквы алфавита, слова, лица и т. д. Когда соответствующий нейрон «узнает» предназначенное ему для узнавания, он возбуждается, и это является физиологическим механизмом восприятия. При повреждении ассоциативных участков развиваются зрительные агнозии (рис.).
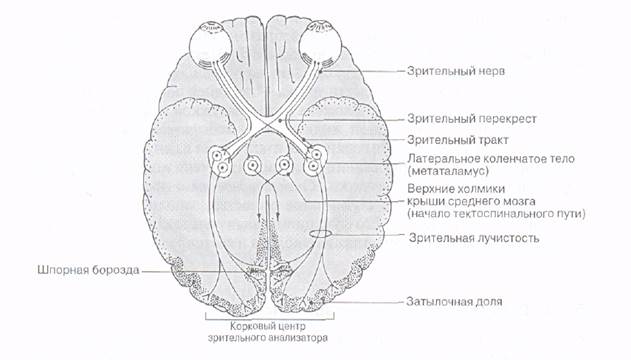
Рис. Проводящий путь зрительного анализатора
Электроретинограмма. При действии света в фоторецепторах и в нейронах сетчатки генерируются электрические потенциалы. Суммарный электрический ответ сетчатки на действие света называют электроретино-граммой (ЭРГ). Она может быть зарегистрирована от целого глаза (для этого один электрод размещают на поверхность роговой оболочки, а другой - на кожу лица вблизи глаза либо на мочку уха) или непосредственно от сетчатки.
На электроретинограмме различают характерные волны - а, b, с, d. Волна а отражает возбуждение внутренних сегментов фоторецепторов (поздний рецепторный потенциал) и горизонтальных клеток. Волна b возникает в результате активации глиальных (мюллеровских) клеток сетчатки ионами калия, выделяющимися при возбуждении биполярных и амакриновых нейронов. Волна с отражает активацию клеток пигментного эпителия, а волна d - активность фоторецепторов и биполярных клеток при выключении света. Следует подчеркнуть, что электрическая активность ганглиозных клеток сетчатки не отражается на ЭРГ.
В целом, ЭРГ хорошо отражает интенсивность, цвет, размер и длительность действия светового раздражителя: амплитуда всех волн ЭРГ увеличивается пропорционально логарифму силы света и времени, в течение которого глаз находился в темноте; волна d (реакция на выключение) тем больше, чем дольше действовал свет. Поэтому ЭРГ широко используется в клинике глазных болезней для диагностики и контроля лечения при различных заболеваниях сетчатки.
Зрительные вызванные потенциалы. Для оценки передачи сигналов на разных уровнях зрительной системы часто используют регистрацию суммарных зрительных вызванных потенциалов (ЗВП), которые у животных можно одновременно отводить от всех отделов мозга, а у человека - от зрительной зоны коры с помощью наложенных на кожу головы электродов. ЗВП регистрируются при предъявлении зрительной стимуляции - вспышек или каких-либо структурированных изображений (шахматной доски, геометрических фигур). В ЗВП, зарегистрированных со скальпа, выделяют волны Р40, N70,Р100-130,а также комплекс волн N240-420. Волны с латентным периодом до 70 мс более выражены во фронтальных отведениях, что связывают со вкладом в ЗВП потенциала электроретинограммы. Начиная с 70 мс волны максимально представлены в постцентральных отведениях - теменных (темпоральных) и затылочных (окципитальных).
В ЗВП (как и в других вызванных потенциалах) выделяют два типа компонентов: ранние специфические (экзогенные) и поздние неспецифичес-кие (эндогенные). Экзогенные компоненты ЗВП регистрируются в интервале до 100 мс от начала ответа; они отражают процессы обработки зрительной информации в сетчатке и подкорковых структурах. Эндогенные компоненты ЗВП, возникающие позднее и длящиеся до окончания ответа, отражают этапы более сложной обработки стимула: формирование образа, сличение его с эталонами памяти, принятие перцептивного решения. Не исключено, что эндогенные компоненты ЗВП отражают процессы параллельного функционирования подсистем, в которых переработка информации о разных признаках стимула происходит независимо, например, по одному каналу осуществляется описание формы объекта, по другому - контраста, цвета и т. д.
Сравнение ЭРГ и ЗВП позволяет установить локализацию патологичес-кого процесса в зрительной системе человека.
Темновая и световая адаптации. Темновая адаптация, т.е. значительное повышение чувствительности глаза наблюдается при переходе из светлого помещения в темное. В первые десять минут пребывания в темноте чувствительность глаза к свету увеличивается в десятки раз, а затем в течение часа - в десятки тысяч раз. В основе темновой адаптации лежат два основных процесса - восстановление зрительных пигментов и увеличение площади рецептивного поля. В первое время происходит восстановление зрительных пигментов колбочек, что, однако, не приводит к большим изменениям чувствительности глаза, так как абсолютная чувствительность колбочкового аппарата невелика. К концу первого часа пребывания в темноте восстанавливается родопсин палочек, что в 100000-200000 раз повышает чувствительность палочек к свету (и, следовательно, повышает периферичес-кое зрение). Кроме того, в темноте вследствие ослабления или снятия латерального торможения (в этом процессе принимают участие нейроны подкорковых и корковых центров зрения), существенно увеличивается площадь возбудительного центра рецептивного поля ганглиозной клетки (при этом возрастает конвергенция фоторецепторов на биполярные нейроны, а биполярных нейронов - на ганглиозную клетку). В результате этих событий за счет пространственной суммации на периферии сетчатки световая чувствительность в темноте возрастает, но при этом снижается острота зрения. Активация симпатической нервной системы и рост продукции катехоламинов повышают скорость темновой адаптации.
При переходе от темноты к свету наступает временное ослепление, затем чувствительность глаза постепенно снижается, т.е. происходит световая адаптация. Она связана, главным образом, с уменьшением площади рецептивных полей сетчатки.
Слепящая яркость света. Слишком яркий свет вызывает неприятное ощущение ослепления. Верхняя граница слепящей яркости зависит от адаптации глаза: чем дольше была темновая адаптация, тем меньшая яркость света вызывает ослепление. Если в поле зрения попадают очень яркие (слепящие) объекты, они ухудшают различение сигналов в значительной части сетчатки (на ночной дороге водителей ослепляют фары встречных машин). При тонких зрительных работах (длительное чтение, сборка мелких деталей, работа хирурга) надо пользоваться только рассеянным светом, не ослепляющим глаза.
|
|
|
|
|
Дата добавления: 2017-01-14; Просмотров: 4735; Нарушение авторских прав?; Мы поможем в написании вашей работы!