КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Вестибулярная сенсорная система
|
|
|
|
Все живые организмы развивались и живут в условиях действия на них силы тяжести, или гравитационного поля Земли, которое практически постоянно по величине и направлению к поверхности Земли и действует на любое тело на Земле. В таких условиях живые организмы на основе оценки направления действия гравитационного поля (т.е. определения «верха», «низа», величины отклонения положения тела от нормального) должны уметь принимать определенное положение по отношению к линии действия силы тяжести, а также уметь координировать свою двигательную активность, при которой нередко требуется значительное отклонение положения тела от нормального. Для оценки направления действия гравитационного поля, т.е. для определения положения организма в трехмерном пространстве и возник вестибулярный (статокинетический) анализатор, или орган статического и динамического равновесия. У человека его периферический отдел представлен вестибулярным аппаратом, или вестибулярным (преддверным) лабиринтом.
Аппарат расположен в лабиринте пирамиды височной кости. В нем различают костную и перепончатую части. Внутри последней находится эндолимфа (ее вязкость в 2-3 раза выше воды), а между костным и перепончатым лабиринтом находится перилимфа. Костный и заполняющий его перепончатый лабиринты состоят из преддверия, трех полукружных каналов и улитки, которая не имеет отношения к органу равновесия. Преддверие (vestibulum, вестибулюм) представляет собой овальную полость, в которой содержится два мешочка - эллиптический (маточка, utriculus) и сферический (собственно мешочек, sacculus), играющие важную роль в оценке положения тела в пространстве. Полукружные каналы располагаются в трех взаимно перпендикулярных плоскостях: передний канал - в сагиттальной плоскости, задний - во фронтальной, а латеральный - в горизонтальной. Такое расположение позволяет осуществлять контроль за различными поворотами головы в любой плоскости. Внутри полукружных каналов залегают одноименные полукружные перепончатые протоки. Одна из ножек каждого перепончатого протока, как и костного, расширяясь, образует перепончатую ампулу (переднюю, заднюю и латеральную). Оба конца каждого полукружного протока открываются в утрикулюс. Соседние ножки переднего и заднего каналов соединяются, образуя общую костную ножку, поэтому три канала открываются в утрикулюс пятью отверстиями.
Участки мешочков, содержащие чувствительные к гравитации и к линейным ускорениям клетки, называются пятнами, или макулами. Аналогичные участки ампул полукружных перепончатых каналов, содержащие клетки, чувствительные к угловым ускорениям или замедлениям, называются гребешками, или кристами.
Вестибулярный анализатор предназначен для пространственной ориентации человека (в том числе при активных и пассивных движениях) по отношению к гравитационному полю Земли, для оценки основных характеристик гравитационного поля, для определения положения головы и шеи по отношению к туловищу, для восприятия линейных и угловых ускорений или замедлений, а также для сохранения равновесия. На основе полученной информации от вестибулярного аппарата при пассивном движении корковые отделы вестибулярного анализатора запоминают направление движения, повороты и пройденное расстояние. При активных движениях вестибулярная система получает, передает и анализирует информацию об ускорениях или замедлениях, возникающих в процессе прямолинейного или вращательного движения, а также при изменении положения головы в пространстве. При равномерном движении или в условиях покоя рецепторы вестибулярной сенсорной системы не возбуждаются. В целом, вся информация, идущая от вестибулярного аппарата в мозг, используется, прежде всего, для регуляции позы и локомоций, т.е. в управлении скелетной мускулатурой.
Следует отметить, что вопрос о значении вестибулярной системы в пространственной ориентировке животных и человека обсуждается в течение длительного времени в связи с гипотезой Циона (1879) о вестибулярном аппарате как анализаторе пространства. В настоящее время существует несколько точек зрения о механизмах ориентации в пространстве, общей чертой которых является представление о комплексном участии ряда сенсорных систем в осуществлении этой функции. Согласно одним представлениям, высшие млекопитающие животные и человек ориентируются в пространстве в основном с помощью зрения и лабиринтов. По мнению других, пространственный анализ представляет собой комплексную функцию динамического ансамбля многих анализаторов, деятельность которых интегрируется двигательным анализатором. Третьи рассматривают ориентацию организма в пространстве как процесс автоматического управления. Задача рецепторов лабиринта в этом процессе заключается в слежении за изменениями в пространстве положения анатомо-физиологического центра черепа, а не всего тела в целом. Информация об изменениях в пространстве положения этого центра, поступающая от вестибулярного аппарата, обогащается информацией других модальностей - зрительной, слуховой, а также информацией о рефлексах положения.
Таким образом, все теории сходятся на представлении о комплексном участии сенсорных систем при ориентации в пространстве с непременным участием вестибулярного аппарата как необходимого условия для формирования адекватного поведения человека и животных в окружающей среде.
Чувствительность вестибулярной системы здорового человека очень высока: отолитовый аппарат дает возможность воспринять ускорение прямолинейного движения, равное всего 2 см/с2. Порог различения наклона головы в сторону - всего около 1°, а вперед и назад - 1,5-2°. Кристы полукружных каналов позволяет человеку замечать ускорения вращения равное 2-3 см/с2.
Общие принципы функционирования вестибулярного аппарата. Для ориентации по отношению к гравитационному полю Земли большинство животных имеет датчик положения тела, т.е. специальный отолитовый орган, построенный по одному и тому же принципу. Орган равновесия, или гравирецептор, состоит из двух частей: «пробной массы», т.е. отолита, обладающего большей или меньшей свободой перемещения в пределах органа, и системы рецепторов, воспринимающих положение или перемещение этой массы в органе. Любое отклонение в положении тела сопровождается смещением пробной массы (отолита, отоконий, отолитовой мембраны), возбуждающей соответствующую группу рецепторов органа равновесия. Сигналы этих рецепторов обрабатываются ЦНС, которая и посылает командный сигнал к мышцам, корректирующим положение тела.
Структуры, ответственные за восприятие угловых ускорений, представляют собой каналы, заполненные жидкостью, в стенках которых располагаются рецепторные клетки. Когда животное или человек перемещается в пространстве, жидкость в канале приходит в движение и отклоняет реснички клеток в направлении, которое зависит от направления ускорения. У человека, как и у высших млекопитающих животных, вестибулярный аппарат содержит одновременно и гравирецепторы, и рецепторы ускорения.
Эволюционные аспекты развития гравитационной сенсорной системы. У многих беспозвоночных (за исключением насекомых) универсальным органом восприятия гравитационного поля является статоцист - замкнутая полость, заполненная жидкостью и содержащая статолит или статоконии (секреты стаоциста или привнесенные из окружающей среды; например, у омаров и креветок - это песчинки). Полость статоциста выстлана реснитчатыми клетками, которые у одних животных расположены равномерно, у других - образуют скопления, называемые макулами. При изменении положения тела происходит смещение статокониев в статоцисте, которое, в свою очередь, вызывает отклонение ресничек рецепторных клеток. Рецепторные клетки статоциста беспозовочных, в отличие от типичных волосковых клеток позвоночных, являются первичночувствующими и имеют центральные отростки. Аксоны рецепторных клеток образуют статический нерв, оканчивающийся в церебральных ганглиях.
В статоцистах головоногих моллюсков, которые занимают в эволюции более высокое положение, наряду с макулами, выполняющими функцию гравитационных рецепторов и обеспечивающими статическое равновесие, имеются и кристы (группы волосковых клеток, покрытые купулами) - рецепторы угловых ускорений, обеспечивающие динамическое равновесие. Таким образом, с развитием двигательной активности животных статоцист, возникший как орган чисто статического равновесия, превращается в орган, обладающий способностью регуляции не только поведения организма в гравитационном поле, но и координации его движений в трехмерном пространстве. Важно также подчеркнуть, что уже у беспозвоночных животных система статоциста функционально связана с другими сенсорными системами, например зрительной, хеморецепторной, с системой рецепторов растяжения, что обеспечивает координацию двигательных актов, регуляцию мышечного тонуса и адаптивное поведение. Наряду с гравитационной функцией система статоциста обладает способностью к сейсмо- и виброрецепции, а у некоторых высокоорганизованных форм - и к акустическому приему. Это позволяет рассматривать сенсорную систему статоциста ракообразных и моллюсков в качестве аналога акустико-вестибулярных систем позвоночных животных.
У насекомых вместо статоцистов функцию рецепторов гравитации выполняют наружные проприоцепторы - волосковые пластинки, а функцию рецепторов угловых ускорений - жужжальца, которые напоминают гироскопы, предназначенные для стабилизации самолетов и подводных лодок.
У позвоночных животных происходит дальнейшее совершенствование вестибулярной системы, особенно ее центрального отдела. Рецепторы гравитации у всех позвоночных животных и человека располагаются в лабиринте. Он представлен двумя (у миноги) и тремя (у всех остальных позвоночных) полукружными каналами, утрикулюсом (маточкой), саккулюсом (мешочком) и лагеной. У высших позвоночных в нем появляется улитка как вырост саккулюса. Верхняя часть лабиринта (полукружные каналы и утрикулюс с их сенсорными элементами) выполняет в основном гравитационные функции (отолитовый аппарат утрикулюса как датчик равновесия, полукружные каналы как датчик угловых ускорений), а нижняя часть (саккулюс, улитка и лагена) - слуховые функции. Поэтому верхнюю часть лабиринта обычно выделяют в качестве преддверия, или вестибулярного аппарата, противопоставляя его органу слуха - улитке.
У всех позвоночных, включая высших млекопитающих животных и человека, рецепция силы тяжести и углового ускорения осуществляется с участием вторично-чувствующих волосковых клеток, возбуждение которых через холинергические синапсы, т.е. с участием ацетилхолина передается на афферентные биполярные нейроны вестибулярного ганглия. Различают колбообразные и цилиндрические волосковые клетки. Оба вида клеток располагаются между поддерживающими (опорными) клетками. Каждая рецепторная, или волосковая, клетка снабжена одной подвижной ресничкой, или киноцилией, и 60-80 склеенными между собой неподвижными волосками, или стереоцилиями. Преобразование действующей силы тяжести, линейного или углового ускорения в рецепторный потенциал осуществляется за счет изменения положения стереоцилий под влиянием отолитовой мембраны (в макулах утрикулюса и саккулюса) или купулы (в кристах полукружных каналов). Это связано с тем, что механическое смещение стереоцилий изменяет проницаемость для ионов (вероятнее всего, для ионов калия). Смещение стереоцилии в сторону киноцилии приводит к деполяризации, т.е. к возбуждению рецептора, а смещение в противоположном направлении вызывает гиперполяризацию, т.е. тормозный эффект. Киноцилия регулирует степень скольжения, давления и точности отхождения на исходные позиции отолитовой мембраны в область каждой ре-цепторной клетки.
У всех млекопитающих макула утрикулюса воспринимает положение организма по отношению к гравитационному полю, т. е. служит рецептором гравитации. Макула саккулюса (кроме рыб) является вспомогательной по отношению к утрикулюсу и, кроме того, хорошо реагирует на вибрацию. Кристы ампул трех полукружных каналов воспринимают угловые ускорения.
На морских свинках показано, что площадь утрикулярной макулы составляет всего 0,55 мм2 и ней содержится примерно 9000 волосковых клеток, для сакку-лярной макулы эти значения составляют 0,50 мм2 и 7500 клеток, а для каждой кристы латерального, переднего и заднего каналов - соответственно 0,40 мм2 и 5500 клеток.
В отолитовых частях лабиринта, т. е. в утрикулюсе и саккулюсе волосковые клетки собраны в виде возвышений, или макул (macula sacculi, macula utriculi). Киноцилии этих клеток либо прикреплены к крупному отолиту (рыбы), либо пронизывают толстую отолитовую мембрану (остальные позвоночные), содержащую кристаллики шестиугольной формы из углекислого кальция (СаСОз); эти кристаллики называют отокониями, или отолитами, или статолитами; все кристаллы склеены между собой желеобразным веществом. Таким образом, отолитовая мембрана представляет собой студенистую, войлокообразную массу, покрывающую всю макулу. Дистальные концы стереоцилий волосковых клеток макулы свободно подпирают или «зачесывают» отолитовую мембрану. Эта мембрана тяжелее остальной ткани и может оказывать давление на волоски рецепторных клеток. Смещение отолитовой мембраны при изменении положения тела в гравитационном поле сопровождается появлением режущих сил, действующих на волоски рецепторных клеток. Тем самым макулы воспринимают изменение силы тяжести и линейного ускорения.
В макулах отолитовых органов ориентация киноцилии такова, что при смещении отолитовой мембраны одни волосковые клетки макулы возбуждаются, другие затормаживаются. В утрикулярной макуле, т.е. в основном рецепторе гравитации позвоночных, рецепторные клетки со сходной поляризацией собираются в группы, которые в совокупности образуют четырехплановую мозаику. Каждой части этой мозаики, или квадранта, может соответствовать определенная группа мышц, принимающих участие в установке тела и регулируемых функций утрикулярной макулы.
Отведение импульсной активности от волокон нервной веточки, иннервирующей утрикулярную макулу обезьяны, показало, что 70% обследованных единиц возбуждалось при наклонах головы в ипсилатеральную сторону, 30% - при наклонах в контралатеральную сторону. Кроме того, около 50% единиц активировалось при наклонах «носом вверх» и примерно столько же при наклонах в противоположном направлении. Эти результаты хорошо согласуются с особенностями пространственной поляризации волосковых клеток в утрикулярной макуле. С другой стороны, они показывают, что адекватным раздражением для утрикулюса являются наклоны головы (вправо, влево, вверх, вниз). Показано также, что рецепторный аппарат преддверия (утрикулюса и саккулюса) возбуждается при ускоряющемся или замедляющемся прямолинейном движении тела и тряске. Таким образом, отолитовый аппарат воспринимает как раздражения ускорения или замедления прямолинейного движения так и изменения в положении головы, вызывающие смещение отолитовой мембраны.
Перепончатые полукружные каналы. Волосковые клетки полукружных каналов, как уже отмечалось, сконцентрированы только в кристах ампул. Волоски погружены в полупрозрачную студенистую массу - купулу, которая не содержит отолитов и покрывает гребешок. Эндолимфа, заполняющая перепончатый проток полукружных каналов, оказывает равномерное влияние на купулу, если голова находится в покое или если животное движется равномерно и прямолинейно. Если же голова поворачи-вается в сторону, то давление эндолимфы на стенки полукружных каналов меняется. Вследствие инерции эндолимфа в канале смещается и сильнее давит на сторону, противоположную движению. Воспринимающие клетки реагируют на это изменение и посылают в мозг соответствующую информацию. Следовательно, воспринимающие клетки полукружных каналов реагируют на угловое ускорение, то есть на перемену направления движения, если даже скорость его остается постоянной. Они воспринимают также вращательное ускорение и замедление.
Экспериментально показано, что в волосковых клетках кристы латерального (горизонтального) канала киноцилии в пучках стереоцилий ориентированы в сторону утрикулюса, а в клетках переднего (сагиттального) канала - в сторону просвета канала. Такая морфологическая поляризация имеет, по-видимому, отношение к механизму направленной (дирекционной) возбудимости канала. Так, в горизонтальном канале ток эндолимфы вызывает возбуждающий эффект, если он направлен к утрикулюсу, a в сагиттальных - тормозной эффект.
Итак, адекватным раздражением для полукружных каналов являются угловые ускорения и замедления, возникающие при вращении человека или животных в горизонтальной, фронтальной или сагиттальной плоскостях, а также смена направления движения, если даже скорость его остается постоянной.
Центральные вестибулярные пути. Первыми нейронами, воспринимающими возбуждение волосковых клеток вестибулярного аппарата, являются биполярные нейроны, составляющие основу вестибулярного, или преддверного, узла (ганглия Скарпе), который залегает на дне внутреннего слухового прохода. Их дендриты, контактирующие с волосковыми клетками, в ответ на возбуждение этих клеток-рецепторов генерируют потенциалы действия, которые передаются в ЦНС по аксонам. Аксоны биполярных клеток образуют вестибулярную, или преддверную, часть преддверно-улиткового нерва (VIII пара черепно-мозговых нервов). Физиологические исследования показали, что в вестибулярном нерве даже при полном покое наблюдается спонтанная электрическая активность. Частота разрядов в нерве повышается при поворотах головы в одну сторону и тормозится при поворотах в другую (детекция направления движения). Реже частота разрядов повышается или, наоборот, тормозится при любом движении. У 2/3 волокон обнаруживают эффект адаптации (уменьшение частоты разрядов) во время длящегося действия углового ускорения.
Волокна вестибулярной части нерва входят в области мостомозжечкового угла в продолговатый мозг и оканчиваются на нейронах бульбарного вестибулярного комплекса. Эти нейроны являются первым уровнем в ЦНС, где происходит обработка информации о движении или изменении положения тела в пространстве, поступающей от рецепторов лабиринта. В состав вестибулярного комплекса, расположенного в области вестибулярного поля на дне ромбовидной ямки, входят четыре ядра - верхнее (ядро Бехтерева), латеральное (ядро Дейтерса), медиальное (ядро Швальбе) и нижнее (ядро Роллера). Нейроны вестибулярных ядер обладают способностью реагировать на изменение положения конечностей, повороты тела, сигналы от внутренних органов, т.е. осуществлять синтез информации, поступающей из разных источников.
Вестибулярные ядра продолговатого мозга связаны со всеми отделами ЦНС: спинным мозгом, мозжечком, ретикулярной формацией ствола мозга, глазодвигательными ядрами, корой головного мозга, вегетативной нервной системой. Среди этих связей можно выделить пять главных проекционных систем.
1. Вестибулоспинальная система, или вестибулоспинальный путь - это аксоны нейронов вестибулярных ядер, идущие к альфа-мотонейронам спинного мозга в виде двух продольных пучков - медиального и латерального. Волокна медиального продольного пучка контактируют с альфа-мотонейронами, управляющими мускулатурой шеи и туловища, а волокна латерального продольного пучка, которые начинаются, главным образом, от латерального ядра Дейтерса, контактируют с альфа-мотонейронами мышц конечностей. Благодаря вестибулоспинальному тракту вестибулярный аппарат участвует в регуляции мышечной активности (за счет реализации статических и статокинетических рефлексов) и тем самым способствует сохранению равновесия в условиях покоя и при движении.
2. Вестибуломозжечковая система, или вестибуло-мозжечковый путь, связывает вестибулярные ядра (через нижние ножки мозжечка) с ядрами шатра мозжечка, т.е. с архиоцеребеллумом, доставляя к нему информацию о положении тела в пространстве. Важно отметить, что к мозжечку информация от вестибулярного аппарата поступает и по прямому сенсорному мозжечковому пути, который начинается непосредственно от нейронов вестибулярного ганглия и идет транзитом до мозжечка через вестибулярные ядра, не прерываясь в них. Все это отражает тесное взаимодействие между вестибулярной сенсорной системой и мозжечком в тонкой координации и регуляции движений. Нарушение связи вестибулярного аппарата и мозжечка может приводить к развитию вестибулярной атаксии, т.е. к нарушению координации движений, что, например, наблюдается при болезни Миньера.
3. Вестибулоокулярная система, или вестибулоокулярный путь, связывает вестибулярные ядра продолговатого мозга с альфа-мотонейронам глазодвигательных нервов (III, IV и VI пары черепно-мозговых нервов). Этот путь идет в составе медиального продольного пучка, который соединяет между собой все ядра глазодвигательных нервов. Эти связи играют важную роль в регуляции движений глаз, что необходимо для сохранения стабильного изображения на сетчатке во время движения. При возбуждении вестибулярных ядер возникают вестибуло-глазодвигательные рефлексы (нистагм глаз, нистагм головы), т.е. ритмические движения глаз и головы в противоположную вращению сторону, которые сменяются скачком глаз и головы обратно. Возникновение и характеристика вращательного нистагма глаз являются важными показателями состояния вестибулярной системы, что широко используется в клинической практике.
Таким образом, нейроны вестибулярных ядер обеспечивают контроль и управление различными двигательными реакциями, в том числе осуществляют динамическое перераспределение тонуса скелетной мускулатуры и включают рефлекторные реакции, необходимые для сохранения равновесия. Мозжечок при этом ответственен за фазический характер этих реакций: после его удаления вестибулоспинальные влияния становятся по преимуществу тоническими. Во время произвольных движений вестибулярные влияния на спинной мозг ослабляются.
4. Вестибуло-гипоталамическая система, или вестибуло-гипоталамический путь, связывает вестибулярные ядра с парасимпатическими (трофотропными) нейронами гипоталамуса, находящимися, главным образом, в преоптической и передней области гипоталамуса. При возбуждении вестибулярных ядер повышается тонус парасимпатической нервной системы, в том числе тонус блуждающего и языкоглоточного нервов, что вызывает при выраженном раздражении вестибулярного аппарата вестибуло-вегетативные реакции - тошноту, рвоту, падение артериального давления, а также кинетоз, т.е. укачивание. Это свидетельствует о том, что за счет повышения тонуса парасимпатической нервной системы вестибулярные ядра (в пределах физиологической нормы) могут принимать участие в регуляции деятельности сердечно-сосудистой системы, желудочно-кишечного тракта и других органов
Однако при сильных и длительных нагрузках на вестибулярный аппарат возникает патологический симптомокомплекс, названный болезнью движения, например, морская болезнь. Болезнь проявляется изменением сердечного ритма (учащение, а затем замедление), сужением, а затем расширением сосудов, усилением сокращений желудка, головокружением, тошнотой и рвотой. Повышенная склонность к болезни движения может быть уменьшена специальной тренировкой (вращение, качели) и применением ряда лекарственных средств.
5. Лемнисковый путь - это путь, по которому информация от вестибулярных ядер поступает (после перекреста на уровне трапециевидного тела) через медиальную петлю (лемниск) к специфическим ядрам таламуса. Вероятнее всего, что у человека информация попадает к нейронам вентробазального комплекса таламуса (в вентральное постлатеральное ядро, в том числе в его дорсомедиальную часть), а от этого ядра начинается прямой путь в кору больших полушарий. Кроме того, предполагается, что существует еще один путь к коре больших полушарий - так называемый непрямой путь - вестибу-лоцеребелло-таламический, который проходит к коре больших полушарий через медиальную часть вентролатерального ядра таламуса.
Обработка информации от вестибулярного аппарата в коре больших полушарий. Вестибулярные проекционные зоны в коре мозга, видимо, различаются у разных представителей млекопитающих. У кошки электрические стимулы, наносимые на вестибулярный нерв или вестибулярные ядра, сопровождаются генерацией потенциалов в передних участках супрасильвиевой и эктосильвиевой извилин височной коры обоих полушарий. Наряду с основной проекционной зоной, находящейся в передней части височной коры, имеется и вторая вестибулярная зона, расположенная каудальнее первой.
У приматов основная вестибулярная зона локализована в задней части постцентральной извилины между первой и второй соматосенсорными зонами. Вторая вестибулярная зона локализуется в моторной коре впереди от нижнего участка центральной борозды. Локализация вестибулярной зоны у человека окончательно не выяснена. Возможно, что у человека вестибулярная зона локализована в нижней области постцентральной извилины, в верхней и средней височных извилинах (по У.Пенфильду), а также в лобных долях, где в моторной зоне (спереди от нижней части центральной борозды) обнаружена вторая вестибулярная зона.
При возбуждении вестибулярного аппарата часто возникают вестибулосенсорные рефлексы, проявляющиеся в головокружении, что представляет собой результат нарушения восприятия пространства (рис.).
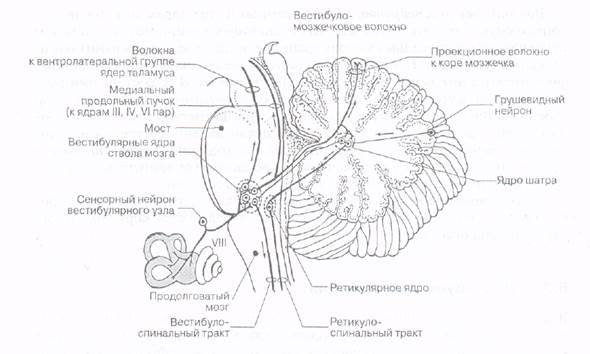
Рис. Проводящий путь вестибулярного анализатора
Адаптация вестибулярного аппарата. У людей ряда профессий (монтажники-высотники, моряки, летчики, космонавты и другие) вестибулярный аппарат подвергается частому и выраженному возбуждению, что приводит к развитию нежелательных вестибуловегетативных, вестибуло-моторных и вестибулосенсорных реакций. Для снижения чувствительности вестибулярного аппарата к действию адекватных раздражителей проводятся тренировки, основу которых составляют процессы торможения реализации указанных реакций. Важным элементом профессионального подбора (в отношении указанных выше профессий) является отбор лиц, обладающих сравнительно высоким порогом возбуждения вестибуловегетативных, вести-було-моторных и вестибулосенсорных реакций.
Развитие космонавтики сделало возможным длительное пребывание человека в космосе, когда невесомость ставит отолитовую систему в совершенно необычные условия. Поэтому изучение реакций отолитовых рецепторов в невесомости представляет первостепенный теоретический и прикладной интерес. При космических полетах в условиях невесомости сила притяжения земли уравновешена центробежной силой, направленной от земли. Невесомым становится весь организм человека, в том числе и его отолиты. Невесомые отолиты перестают давить на рецепторные клетки, и мозг не получает информации о том, где верх и низ. Отолиты отстают от движения головы и при резких ее поворотах вызывают необычные раздражения чувствительных клеток, что ведет к сильной головной боли, рвоте и пр.
Однако, по свидетельству космонавтов, к невесомости они привыкают быстро, в течение всего нескольких дней. Такая способность к адаптации свидетельствует о необыкновенной пластичности всей нервной системы, если учесть, что вестибулярная система эволюционировала в течение миллионов лет, никогда не попадая в подобные условия.
Методы исследования вестибулярного анализатора. Состояние вестибулярного аппарата оценивается с помощью различных функциональных проб, в том числе вращательной пробы, пробы Ромберга, пробы на походку, пробы на нистагм.
Вращательная проба предназначена для адекватного раздражения ампулярного отдела вестибулярного анализатора путем вращения тела вокруг вертикальной оси на специально созданных для этих целей креслах, например, на кресле Р.Барани. Проба позволяет оценить состояние
вестибулярного анализатора по появлению вестибулосенсорных (головокружение), вестибуловегетативных (побледнение лица, потоотделение на лице) и вестибуломоторных реакций (нистагм головы и глаз), а также судит о возбудимости ампулярного отдела по наименьшей угле вой скорости (т.е. по углу поворота за 1 с), вызывающей соответствующие реакции. У здорового человека вестибулосенсорные реакции, например, ощущение противовращения, возникают при пороговой скорости, равной 0,5°- 4,0° в 1 с. Для выполнения пробы с целью изучения вестибуломоторных реакций испытуемого усаживают в кресло Р.Барани, закрывают переднюю перекладину кресла производят вращение со скоростью 0,5 оборотов за 1 с (это превышает пороговую в 100 раз). После 10 оборотов кресло внезапно останавливают. При нормальном функциональном состоянии лабиринтов наблюдается (обычно в течение 20-30 с) нистагм головы и глаз - они медленно поворачиваются в сторону, противоположную вращению, а затем быстро возвращаются в исходное положение.
Указательная проба в модификации Р.Барани заключается в том, что испытуемый до и после вращения на кресле Р.Барани должен выполнить указательные пробы, например, с закрытыми глазами коснуться кончика ка- рандаша, установленного перед ним на расстоянии 0,5 м на уровне его головы.
Проба Ромберга используется с целью выявления вестибулярной атаксии, т.е. нарушения регуляции двигательной активности (координации движений) вследствие поражения вестибулярного аппарата. Для этого испытуемому предлагается стоять со сдвинутыми ногами и закрытыми глазами. При наличии вестибулярной атаксии больной шатается или падает; эти явления усиливаются при поворотах головы. При отсутствии симптомов атаксии применяют усложненную пробу Ромберга - испытуемому предлагается стоять, выставив одну ногу перед другой, или стоять на пальцах стопы, либо стоять на одной ноге. При наличии вестибулярной атаксии больной шатается или падает, а выключение зрения усиливает эти нарушения.
Проба на походку проводится с целью выявления вестибулярной атаксии. Испытуемому предлагается пройти вперед и назад (по прямой линии) и в стороны (фланговая походка). При вестибулярной атаксии больной шатается или падает, а выключение зрения усиливает эти нарушения.
Проба на нистагм также проводится с целью выявления вестибулярной атаксии. Испытуемый должен, не поворачивая головы, смотреть вверх, вниз, вправо, влево с фиксацией взора на пальце исследователя. При вестибулярной атаксии могут наблюдаться разные виды нистагма глаз - горизонтальный, вертикальный, ротационный. Возникновение и характеристика вращательного глазного нистагма являются важными показателями состояния вестибулярной системы, их широко используют в авиационной, морской и космической медицине, а также в экспериментах и клинике.
Соматическая сенсорная система (тактильная, ноцицептивная и температурная сенсорные системы)
Соматическая сенсорная система - это система, предназначенная для анализа механических, химических и температурных воздействий окружающей среды на кожные и слизистые покровы организма человека и животных. Этой системой осуществляется три вида рецепции (или чувствительности) - болевая (ноцицептивная), сигнализирующая о возможности повреждения организма, температурная (терморецептивная), отражающая температуру окружающей среды, и тактильная (механоре-цептивная), отражающая воздействие на кожу и слизистые оболочки различных механических стимулов, вызывающих чувство прикосновения, давления или вибрации. Соответственно этим трем видам рецепции соматическая сенсорная система условно подразделяется на три самостоятельные сенсорные системы - ноцицептивную, температурную и тактильную (ноцицептивный, температурный и тактильный анализаторы). Основная часть периферических отделов соматической сенсорной системы расположена в коже. Поэтому кожу называют органом чувства осязания, температуры и боли, а сама соматическая сенсорная система иногда называется кожным анализатором. Осязанием называют весь комплекс ощущений, возникающих при соприкосновении кожи с различными телами. Некоторые авторы предлагают совокупность соматической (кожной) и проприоцептивной сенсорных систем называть соматосенсорной системой. Однако, по мнению В.И.Циркина и С.И. Трухиной, такое объединение усложняет ориентацию в терминах.
|
|
|
|
|
Дата добавления: 2017-01-14; Просмотров: 1228; Нарушение авторских прав?; Мы поможем в написании вашей работы!