КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Генетические исследования деятельности сердечно-сосудистой системы 1 страница
|
|
|
|
Кровяное давление. Оценки наследуемости кровяного давления, приводимые в разных работах, колеблются от 13% до 82% для систолического давления и от 0% до 64% для диастолического давления (цит. по Boomsma D., 1992), составляя в среднем 50% (Hopkins P., Williams R., 1989); при этом наследуемость показателей давления часто зависит от пола испытуемых. (Mcllhany M. et al., 1975; Schieken R. et al., 1989) и возраста (Province M., Rao D., 1985; Sims J. et al., 1986). Симс с соавторами (Sims J. et al., 1986) обнаружили уменьшение влияния наследственных факторов на уровень диастолического давления у взрослых при переходе от молодого к зрелому возрасту (соответственно 68% и 38%) за счет увеличения влияния несистематической среды. Оценки наследуемости уровня кровяного давления в исследованиях семей, где родственники различаются по возрасту, обычно ниже, чем в близнецовых исследованиях, где оба близнеца имеют одинаковый возраст. В первом случае они составляют 19—45% для систолического и 15—52% для диастолического давления (Iselius L. et al., 1983; Rice Т. et al., 1989), во втором — 41—82% и 51—66% соответственно (Mcllhany M. et al., 1975; Bourhani N. et al., 1976; Slattery M. et al., 1988; Hunt S. et al., 1989;
Schieken R. et al., 1989).
Результаты исследования половых различий в уровне наследственной обусловленности кровяного давления в состоянии покоя неоднозначны. Макилани и соавторы (Mcllhany M. et al., 1975) при обследовании 200 пар близнецов в возрасте около 14 лет обнаружили больший вклад генотипа в вариативность систолического и диастолического давления у девушек, по сравнению с юношами (в группе девушек — 78% и 61%, юношей — 41% и 56% соответственно). Шикен с соавторами (Schieken R. et al., 1989), обследовав выборку из 251 пары близнецов в возрасте 11 лет, не обнаружили половых различий в уровне наследуемости систолического давления (66%). Для диастолического давления наблюдалось несколько большее влияние генотипа в группе мальчиков (64%), по сравнению с девочками (51%). Тамбс с соавторами (Tambs R. et al., 1992) не обнаружили половых различий в степени генетического контроля уровня давления на очень большой выборке испытуемых, составляющей около 75000 родственников различной степени родства.
К настоящему времени накоплено огромное количество фактов, свидетельствующих о ритмичности большинства физиологических функций. В частности, Е.И.Заславской и А.П.Великоиваненко выявлен ритм суточных колебаний показателей кровообращения: в полуденное и вечернее время наблюдается повышение систолического, среднего и пульсового давления, ночью — снижение этих показателей;
значимых колебаний диастолического АД в течение суток не наблюдается; венозное давление существенно повышается в послеполуденные часы и мало изменяется в остальное время суток. Эти данные очень важны с практической точки зрения, поскольку клинические наблюдения свидетельствуют о большей частоте нарушений ритма сердечной деятельности в ночное время.
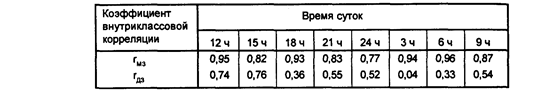
Интересное исследование на близнецах было проведено Р.М.Заславской и И.Х.Олевским (1980). Для генетического анализа суточного ритма кровообращения авторы использовали данные 7 пар МЗ и 6 пар ДЗ близнецов. Определение параметров кровообращения проводили 8 раз в сутки через каждые 3 часа. Измеряли систолическое и диастолическое АД, среднее и пульсовое давление, ударный и минутный объем сердца, ЭКГ. В таблице приведены коэффициенты внутриклассовых корреляций в парах МЗ и ДЗ близнецов. Можно видеть, что наблюдается определенная динамика в уровне внутрипарного сходства. Можно предположить, что уровни систолического Давления ночью, утром и в полдень испытывают сильные генетические влияния. На рисунке 2 приведены графики суточной динамики систолического АД у МЗ и ДЗ близнецов.
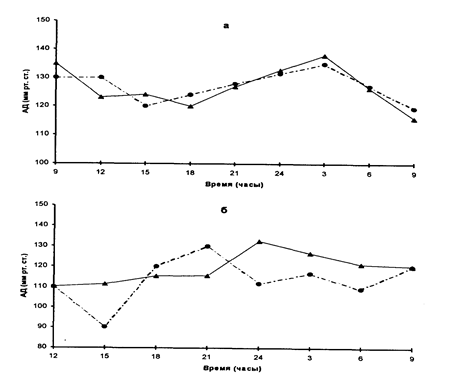
Рисунок 2
Суточная динамика систолического АД у монозиготных (а) и дизиготных (б) близнецов
Аналогичные закономерности наблюдаются для среднего артериального давления. Авторы считают, что максимально сниженное систолическое, среднее и пульсовое АД в ночные и утренние часы (базальный уровень) генетически детерминировано и сложилось в ходе эволюции. Повышение АД, наблюдаемое обычно в послеполуденное время, определяется средовыми факторами, воздействие которых осуществляется благодаря увеличению продукции гормонов коры надпочечников.
Таблица 1
Внутриклассовые корреляции показателей систолического АД в течение суток
Особый интерес представляют исследования природы вариативности реактивных изменений деятельности сердечно-сосудистой системы под влиянием психологических нагрузок. Это дает возможность оценить влияние наследственных факторов на процессы адаптации. В ходе эволюции было выработано множество приспособительных реакций организма на специфические раздражители внешней среды, в том числе такая общая реакция, как стресс. Стресс, или адаптационный синдром, — стандартная неспецифическая реакция организма на чрезмерное раздражение. При стрессе усиливается деятельность адаптационных механизмов, обеспечивающих жизнеспособность индивида в изменившихся условиях внешней среды. Выделяют два типа стрессоров — физические и эмоциональные. При действии физических стрессоров наступает непосредственное нарушение гомеостаза; в случае эмоциональных раздражителей нет действия собственно повреждающих факторов, но есть угроза повреждения. Стресс возникает только тогда, когда сложившаяся ситуация оценивается как опасная. Е.И.Соколов и его коллеги (1980) провели специальное генетическое исследование изменения деятельности сердечно-сосудистой системы в ответ на эмоциональные нагрузки. В исследовании приняли участие 24 пары монозиготных и 15 пар дизиготных близнецов. В качестве эмоциональной нагрузки использовали ситуацию решения интеллектуальной задачи в условиях дефицита времени и воздействия раздражающих стимулов (свет, звук). У всех близнецов в состоянии покоя перед нагрузкой, в процессе решения задач (в начале, в середине и в конце) и через 10 минут после нагрузки измеряли артериальное давление (АД), ЧСС, записывали реоэнцефалограмму (РЭГ) (РЭГ - запись динамики кровенаполнения сосудов головного мозга).
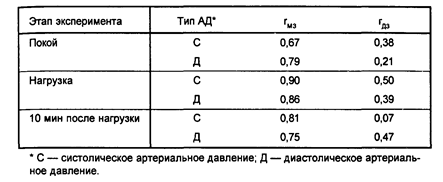
Результаты свидетельствуют о значительном влиянии генетических факторов как на систолическое, так и на диастолическое АД в состоянии покоя, причем для диастолического АД оно выражено сильнее. При нагрузке влияние генетических факторов на систолическое АД резко возрастает, тогда как для диастолического АД остается на прежнем уровне. Увеличение при нагрузке величины коэффициента наследуемости для систолического давления почти в 2 раза позволяет, по мнению авторов, сделать важный вывод:
под действием эмоциональной нагрузки мобилизуются генетически обусловленные адаптационные механизмы.
Когда АД измеряется спустя 10 минут после нагрузки, для систолического АД сохраняется высокий коэффициент наследуемости, тогда как для диастолического он уменьшается (таблица 2).
Таблица 2
Внутриклассовые корреляции показателей артериального давления при эмоциональной нагрузке (по Соколову Е.И. и др., 1980)
При анализе деятельности сердечно-сосудистой системы при физической нагрузке авторами были выявлены похожие закономерности.
Исследование Д.Бумсма с соавторами (Boomsma D. et al., 1992), выполненное на большой выборке близнецов (160 пар в возрасте от 14 до 21 года), показало, что у женщин при нагрузках (задачи на время реакции и решение задач) увеличивалась оценка наследуемости уровня как систолического, так и диастолического давления, в то время как у мужчин данная тенденция наблюдалась только для систолического давления. Диастолическое давление мужчин при нагрузках контролировалось генотипом даже в меньшей степени, чем в покое. Для систолического давления у женщин и мужчин и для диастолического давления у женщин наблюдалось также влияние систематической среды, уменьшающееся при функциональных нагрузках.
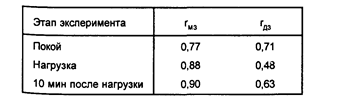
Частота сердечно-сосудистых сокращений (ЧСС). Анализ ЧСС в исследовании Е.И.Соколова и коллег (1980) показал, что в состоянии покоя сходство монозиготных близнецов лишь ненамного превышает сходство дизиготных, тогда как при нагрузке сходство МЗ заметно увеличивается, а сходство дизиготных близнецов уменьшается (таблица 3), что говорит об увеличении коэффициента наследуемости.
Таблица 3
Внутриклассовые корреляции показателей ЧСС при
эмоциональной нагрузке (по Соколову Е.И. и др., 1980)
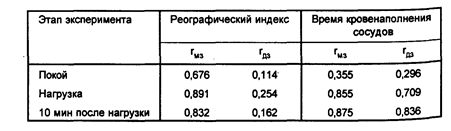
Авторы полагают, что более интенсивное функционирование сердечно-сосудистой системы при нагрузке, компенсируя возрастающие потребности тканей в кислороде, в первую очередь обеспечивает потребности центральной нервной системы. Действительно, уровень кровенаполнения сосудов головного мозга во время эмоционального напряжения несколько возрастает, а во время отдыха снижается. Оказалось, что в состоянии покоя для величин реографического индекса характерен определенный вклад генетических факторов в их вариативность, однако влияние генотипа значительно увеличивается во время нагрузки и уменьшается после 10-минутного отдыха. Анализ времени кровенаполнения сосудов показал, что в состоянии покоя изменчивость этого показателя обусловлена средовыми факторами, а во время нагрузки — наследственными (таблица 4).
Таблица 4
Внутриклассовые корреляции показателей реографического индекса и времени кровенаполнения сосудов (по Соколову Е.И. и др.. 1980)
Сомсен с соавторами (1985) на близнецах исследовали влияние генетических факторов на форму реактивного изменения ЧСС. Фазическое изменение ЧСС представляет собой характерный паттерн, зависящий от конкретной задачи и обладающий высокой межиндивидуальной вариативностью. При этом паттерн реактивности, характерный для индивида, достаточно высоко стабилен. В исследовании Сомсена давались пробы на время реакции (ВР).
Обычно при предъявлении предупреждающего сигнала (S1) происходит замедление ЧСС, затем ускорение, а перед предъявлением "пускового" (S2) сигнала — опять замедление. Ситуация эксперимента представляла собой либо чистые пробы на ВР, либо такие же пробы, перемежающиеся задачами на вычисление в уме. Во втором случае, который можно рассматривать как более "стрессовую" ситуацию, корреляция волновой формы ЧСС-реакции была достоверно выше у МЗ, чем у ДЗ близнецов. В первой ситуации (только задачи на время реакции) различия между коэффициентами корреляции МЗ и ДЗ не были значимы. Авторы считают возможной двоякую интерпретацию этих данных.
Во-первых, более низкая оценка наследуемости ЧСС-реакции в менее стрессовой ситуации может быть обусловлена не собственно меньшей наследуемостью признака, а большей ошибкой измерения, поскольку некоторыми исследователями была показана более высокая стабильность ЧСС-реакции в стрессовых условиях. Во-вторых, различия могут объясняться различной наследуемостью для параметров активности симпатической и парасимпатической систем. Предполагается, что ЧСС в задаче на время реакции определяется в основном активностью парасимпатической системы, а при стрессе (вычисления в уме) — симпатической. Вместе с тем существуют данные о генотипическом контроле биохимических коррелятов активности именно симпатической нервной системы (Propping Р. et al., 1973, цит. по Somsen R. et al., 1985). Возможно, больший вклад генотипа в ситуации перемежающегося предъявления задач на ВР и на вычисление в уме связан с тем, что здесь преобладает активность симпатической вегетативной системы, контролируемой генотипом.
Последнее предположение подтверждается и данными, полученными Б.И.Кочубеем при регистрации ЧСС в ситуациях ориентировочной (ОР) и оборонительной (ОбР) реакций на звук (Кочубей Б.И., 1985, 1988). Для ОР типично преобладание фазы замедления ритма, физиологическим механизмом которого является активность блуждающего нерва (парасимпатическая система), а психологическим коррелятом — "обращенность вовне". Для ОбР, наоборот, характерно симпатически обусловленное ускорение ритма, свидетельствующее об избегании стимула. Стимул 80 дБ вызывал уменьшение ЧСС, а при привыкании ОР наблюдалось относительное учащение ритма. Звук 105 дБ приводил к значительному увеличению ЧСС, а при повторении этого стимула отмечалось относительное снижение ЧСС.
Значительный вклад наследственных факторов фиксировался автором в случае изменения ЧСС в ответ на тон 105 ДБ (оборонительная реакция — ОбР) и отсутствовал для ЧСС-реакции на тон 80 дБ (ориентировочная реакция — ОР). Величина привыкания, наоборот, была обусловлена генотипом только при звуке 80 дБ. Таким образом, генетические влияния наблюдались в ситуациях, в которых ведущую роль играет симпатическая регуляция сердечного ритма (угашение ОР и первая реакция при ОбР), тогда как в ситуациях, характеризующихся парасимпатическим доминированием, вариативность ЧСС определялась исключительно средовыми факторами.
Сходство параметров сердечно-сосудистой деятельности исследовалось также у сибсов (Ditto В., 1987). У 36 пар сибсов в состоянии покоя и при различных функциональных нагрузках измеряли ЧСС, систолическое и диастолическое давление крови, увеличение объема пальца при пульсации (Digital blood volume pulse — DBVP) и время прохождения пульсового импульса до уха (Ear Pulse Transite Time — ЕРТТ). Коэффициенты внутриклассовой корреляции для систолического, диастолического давления и ЧСС в состоянии покоя составляли соответственно 0,62, 0,42 и 0,28. При функциональных нагрузках коэффициенты корреляции были ниже, но для диастолического давления, ЧСС и DBVP достигали значимости в различных условиях.
Д.Бумсма с коллегами (Boomsma D. et al., 1990) провели также исследование респираторной синусной аритмии у близнецов (РСА). РСА отражает циклические изменения ЧСС, сопровождающие дыхание. Частота сердечных сокращений обычно увеличивается при выдохе и уменьшается при вдохе. Чем больше амплитуда изменения ЧСС, тем выше РСА и тем сильнее парасимпатический контроль ЧСС. Выраженный парасимпатический контроль ЧСС и, следовательно, высокая РСА являются признаком хорошего состояния как сердечно-сосудистой системы, так и ЦНС (Porges Р., 1985, цит. по Boomsma D. et al., 1990). В исследовании принимало участие 70 пар МЗ и 80 пар ДЗ близнецов. Как и в предыдущих работах, показатель РСА оценивался в покое и в условиях эмоционального стресса (два типа задач: на время реакции и на вычисления в уме). Генетический анализ данных показал, что при нагрузках около 50% фенотипической изменчивости РСА обусловлено генетическими факторами, тогда как в покое — лишь 25%.
Таким образом, видимо, правы Е.И.Соколов с коллегами, предполагая, что мобилизация при нагрузке генетичес-
ки обусловленных адаптационных механизмов приводит к увеличению показателей наследуемости в этих ситуациях.
Электрокардиограмма (ЭКГ). Генетическим исследованиям ЭКГ посвящены несколько работ. В исследованиях, проведенных в СССР в начале тридцатых годов, было обнаружено, что изменчивость ЭКГ испытывает значительное влияние наследственности (Кабаков И.Б., 1933; Кабаков И.Б., рыбкин И.А., 1934). Эти результаты подтверждаются и в более поздних работах, в которых продемонстрировано значительное внутрипарное сходство характеристик ЭКГ у монозиготных близнецов. В одной из них (рыбкин И.А. и др., 1980) исследование ЭКГ было проведено на достаточно большой выборке близнецов (310 пар), разделенной на три возрастные группы (младшая группа — 16—24 года, средняя 25—44 года, старшая — 45 лет и старше). Сопоставление формы ЭКГ, записанной в 12 отведениях у моно- и дизиготных близнецов, показало, что коэффициент наследуемости основной формы ЭКГ высокий в младшей возрастной группе и уменьшается с возрастом (см. таблицу 5)
Таблица 5
Коэффициенты наследуемости формы ЭКГ в трех возрастных группах
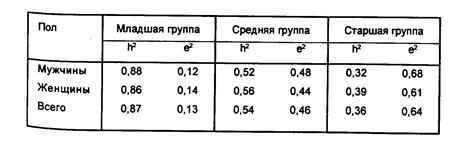
Авторы считают, что как форма ЭКГ в целом, так и ее отдельные элементы в значительной мере обусловлены наследственными факторами, однако с возрастом влияние средовых факторов (особенно у мужчин) возрастает.
В исследовании Соколова Е.И. и коллег (1980) изучалась наследственная обусловленность ЭКГ при физической нагрузке. Полученные результаты свидетельствуют о том, что изменчивость интервала PQ, амплитуда зубцов Р и Т обусловлены генетическими причинами (h2 > 0,5).
Таким образом, результаты исследований сердечно-сосудистой системы свидетельствуют о влиянии наследственных факторов на межиндивидуальную вариативность показателей ее деятельности, причем величина коэффициентов наследуемости зависит от пола и возраста испытуемых, и даже от времени суток, когда регистрируется тот или иной показатель. Так же как и в случае электрокожной активности, влияние наследственных факторов возрастает при различных нагрузках.
Приложение
к лабораторному занятию № 13 «Генотип и среда в индивидуальном развитии»
1. В возрастной психологии рассматривается 4 типа экспериментальных показателей, свидетельствующих о стабильности и преемственности развития — 1) абсолютные величины психологических характеристик, 2) их дисперсии, 3) ковариации (связи) разных показателей, измеренных в одном и том же возрасте, и 4) ковариации показателей, измеренных в разных возрастах.
Рассмотрим, каким образом стабильность и преемственность развития рассматриваются в психогенетике.
Все данные, получаемые в психогенетике носят популяционный характер, то есть все данные, которые получают при сопоставлении различных пар родственников (близнецов, родителей и детей и т.д.), имеют отношение не к абсолютным величинам изучаемых характеристик, а только к широте индивидуальных различий по исследуемой характеристике (или, что то же самое, — к дисперсии исследуемой характеристики, к ее межиндивидуальной вариативности).
Популяционный характер данных о наследственности и среде определяет формулировку основного вопроса, на который психогенетика пытается ответить при изучении развития: в какой степени генотипические и средовые факторы влияют на возрастные изменения индивидуальных различий по той или иной психологической характеристике.
Соответственно этому в психогенетике выделяются три аспекта анализа возрастных изменений, ассимилированные из возрастной психологии. Это, во-первых, изменение с возрастом дисперсий психологических характеристик (и изменение относительного вклада генотипа и среды в эти дисперсии). Во-вторых, изменение ковариации переменных (или изменение корреляционных и факторных структур психологических характеристик, происходящее в процессе развития, и изменение роли генотипа и среды в ковариациях психологических характеристик). И, в-третьих, изменение межвозрастных ковариации, а также генетических и средовых корреляций между психологическими измерениями, полученными в разных возрастах
Рассмотрим, как исследуется в психогенетике поведения каждый из этих трех аспектов возрастных изменений.
1. О возрастных изменениях вклада генотипических и средовых факторов в дисперсии психологических характеристик судят на основании сравнения компонент фенотипической дисперсии, полученных при анализе одной и той же психологической характеристики в разных возрастах. Например, можно сопоставить внутрипарное сходство МЗ и ДЗ близнецов по интеллекту в 10 лет и в 15 лет. Различия генотип-средового соотношения, полученного в разных возрастах, будут свидетельствовать об изменении в процессе развития роли генотипа и среды в детерминации интеллекта (его индивидуальных различий).
2. С возрастом могут меняться генотипические и средовые причины ковариации переменных. Если в психологическом исследовании обнаружены связи между двумя характеристиками, каждая из которых испытывает влияние генотипа, можно поставить вопрос о том, в какой мере связи между этими характеристиками опосредованы генотипом, а в какой — средовыми факторами.
Для того чтобы ответить на этот вопрос, в психогенетике разработаны специальные методы. Так, например, в близнецовом исследовании о роли генотипа и среды в ковариации двух переменных (например, вербального и невербального интеллекта) можно судить по кросс-корреляциям одного и другого близнеца. Для этого надо случайным образом разделить выборку МЗ близнецов так, чтобы в каждую выборку вошел один близнец из пары. Подсчитав корреляции между вербальным интеллектом МЗ близнецов, вошедших в одну выборку, и невербальным интеллектом МЗ близнецов, вошедших во вторую выборку, мы получим кросс-корреляции между вербальным и невербальным интеллектом МЗ близнецов. Иначе говоря, мы выясним, насколько вербальный интеллект одного МЗ близнеца в паре похож на невербальный интеллект второго МЗ близнеца в паре.
Выполнив аналогичную процедуру для ДЗ близнецов, мы получим кросс-корреляции для ДЗ близнецов.
Удвоенная разность между показателями кросс-корреляций МЗ и ДЗ близнецов и представляет собой вклад генотипа в фенотипическую корреляцию и математически выражается следующим образом:
2 (r МЗ – r ДЗ) = r G √h12 ∙ h22
где r МЗ и r ДЗ — кросс-корреляции МЗ и ДЗ близнецов по двум характеристикам (в нашем примере — по вербальному и невербальному интеллекту); h12 и h22 — показатели наследуемости для характеристик вербального и невербального интеллекта; r G — генетическая корреляция между показателями вербального и невербального интеллекта.
Такой анализ фенотипических корреляций позволяет понять причины, лежащие в основе связи, наблюдаемой между двумя характеристиками, и определить, в какой степени генетические факторы, а в какой степени — средовые обусловливают связь между двумя характеристиками.
Сопоставление генетических и фенотипических корреляций дает, кроме того, возможность сделать и некоторые выводы о природе генетических влияний. Так, если обнаруживается вклад генотипа в фенотипическую корреляцию между двумя чертами, это означает, что сравниваемые черты испытывают сходные генетические влияния. Если же обе черты имеют высокие показатели наследуемости, но не связаны фенотипически (не коррелируют между собой), это означает, что гены, определяющие дисперсию одной черты, не определяют дисперсию другой черты. Упрощая, можно сказать, что в первом случае на индивидуальные различия, наблюдаемые по одной черте, влияют одни и те же гены, а во втором случае — разные.
Если роль генотипа и среды в ковариации между переменными оказывается одинаковой в разных возрастах, можно говорить о стабильности развития, то есть о том, что по мере взросления не меняются причины, лежащим в основе взаимосвязей двух переменных.
3. По мере развития может меняться роль генотипа и среды в межвозрастных ковариациях. В психогенетике неоднократно предпринимались попытки связать фенотипическую стабильность признака с его генетической обусловленностью. В течение долгого времени они носили чисто описательный характер и исходили из явной параллельности двух процессов — увеличения с возрастом генетической обусловленности, например, показателей интеллекта и увеличения с возрастом его стабильности.
В настоящее время роль генотипических и средовых факторов в возрастной стабильности и в возрастном изменении является предметом рассмотрения в лонгитюдных исследованиях, организованных по правилам генетического исследования (то есть позволяющих сопоставить внутрипарное сходство различных родственников) и дающих возможность количественно оценить вклад генотипа и среды в фенотипическую стабильность психологических характеристик.
Выделение генотипической и средовой компонент в фенотипической стабильности признака производится аналогично разделению на генотипические и средовые компоненты ковариации между разными психологическими чертами. Так, вклад генотипа в фенотипическую стабильность признака, вычисленный на основании близнецовых данных, будет равен:
2 (r МЗ – r ДЗ) = r G √h12 ∙ h22
где r МЗ и r ДЗ — кросс-корреляции МЗ и ДЗ близнецов по одной и той же психологической характеристике, измеренной в первом и во втором возрастах; h12 и h22 — показатели наследуемости для одной и той же характеристики, полученные в первом и втором возрастах; r G — генетическая межвозрастная корреляция.
Этот способ выделения генетических и средовых компонент в фенотипической стабильности психологических характеристик применим для анализа причин (генотипических и средовых источников вариативности) разных возрастных изменений, например, повышения стабильности психологических характеристик с возрастом или снижения фенотипической стабильности во время перестройки психологических характеристик.
Таким образом, при анализе стабильности и преемственности развития в психогенетике рассматривается 3 типа экспериментальных показателей: 1) дисперсии психологических характеристик, 2) ковариации разных показателей, полученных в одном и том же возрасте, и 3) ковариации показателей, полученных в разных возрастах. Оценивая вклад генотипа и среды в дисперсии и ковариации различных психологических показателей, психогенетика выясняет механизмы, лежащие в основе возрастных изменений, и, следовательно, углубляет представления о закономерностях развития, которые получены в возрастной психологии.
2. Диагностика стабильности и преемственности развития осуществляется двумя методами — сравнительно-возрастным (или методом поперечных срезов) и лонгитюдным.
Сравнительно-возрастной метод основан на сравнении групп испытуемых разного возраста. Он позволяет достаточно быстро (в одной экспериментальной сессии) сопоставить разновозрастные выборки и сделать вывод о различиях между ними. К сожалению, выводы таких исследований не всегда можно интерпретировать с точки зрения возрастных различий.
Применение сравнительно-возрастного метода предполагает эквивалентность сравниваемых в эксперименте групп по всем параметрам, кроме возраста. От того, насколько тщательно будет выполнено это условие, зависит, можно ли будет интерпретировать полученные различия между группами как имеющие отношение к возрастным различиям. Например, если сравнить мальчиков 10 лет и девочек 11 лет, то в полученных результатах будут смешаны возрастные и половые различия, и определить, какова роль собственно возрастных различий в полученных результатах, будет невозможно.
Создание однородных выборок — более сложная проблема, чем это может показаться на первый взгляд. Прежде всего, любые психологические характеристики, и тем более закономерности их развития, связаны с огромным количеством переменных. Учесть в исследовании все эти переменные просто нереально. Из-за этого в результатах, полученных методом поперечных срезов, всегда остается вероятность того, что полученные различия связаны не только с возрастными изменениями, но и с различиями в выборках, которые не контролировались в эксперименте и остались неизвестными экспериментатору. Обычно острота этой проблемы снижается с помощью создания больших репрезентативных выборок испытуемых. Часть параметров в сравниваемых выборках уравнивается, а про другие — предполагается, что различия нивелируются из-за большого количества испытуемых.
Вторая проблема сравнительно-возрастного метода, представляющаяся принципиально неразрешимой, связана с принадлежностью испытуемых, одновременно участвующих в эксперименте, к различным когортам (то есть с тем, что группы испытуемых, которые сопоставляются в эксперименте, имеют разные годы рождения).
Для того чтобы понять, почему это важно, представим себе такую экспериментальную работу. Психологи, желая сопоставить возрастные различия, воспользуются архивными данными и возьмут из них результаты диагностики интеллекта 8-летних детей, полученные в 20-ые годы. С помощью методов, которые использовались в те незапамятные времена, они протестируют современных 9-летних детей. Можно ли будет на основании этих данных судить о различиях в интеллекте 8- и 9-летних детей? Разумеется, нет. Сравниваемые группы детей радикальным образом различаются по условиям существования. Дети читают разные книги, занимаются по разным программам в школе, им доступны совершенно различные источники информации, социально-экономические условия жизни детей абсолютно не сопоставимы, и вообще легче сказать, чем эти группы не похожи, чем найти между ними какое-то сходство. Но ведь эти кардинальные различия появились не внезапно, они накапливались понемногу из года в год, и поэтому современные дети (допустим, те, которые родились в 1989 и в 1990 г.) тоже чуть-чуть различаются по условиям жизни. Из-за этого, сравнивая современных 8- и 9-летних детей, мы получим в возрастных различиях небольшие искажения, связанные с различиями в условиях жизни. Чем больше будет возрастной интервал, который исследуется (чем больше различия в возрасте между группами), тем в большей степени возрастные различия окажутся смешанными со спецификой условий развития сравниваемых когорт.
|
|
|
|
|
Дата добавления: 2014-11-08; Просмотров: 766; Нарушение авторских прав?; Мы поможем в написании вашей работы!