Утворення АТФ з АДФ та Фн може відбуватися тільки в певних ділянках електронотранспортного ланцюга мітохондрій, в яких величина хімічної енергії, що виділяється при транспортуванні пари електронів між двома редокс-системами (компонентами дихального ланцюга), достатня для синтезу 1 молекули АТФ (тобто > 7,3 ккал, або 30,5 кДж).
Ділянки дихального ланцюга мітохондрій, де вивільнення хімічної енергії достатнє для синтезу молекули АТФ
Комплекс І (НАДН → коензим Q)
12,2 ккал
51,0 кДж
Комплекс III (цитохром b → цитохром с1)
9,9 ккал
41,4 кДж
Комплекс IV (цитохром а3 → О2)
23,8 ккал
99,6 кДж
Зазначені ділянки електронотранспортного ланцюга називаються пунктами спряження дихання (електронного транспорту) з окисним фосфоритуванням.
35. Хеміосмотична теорія окисного фосфорилювання, АТФ-синтетаза мітохондрій.
Хеміосмотична теорія передбачає, що:
1. Функціонування дихального (електронотранспортного) ланцюга у внутрішніх (спрягаючих) мембранах мітохондрій супроводжується генерацією на цих мембранах електрохімічного градієнта протонів (Н+).
2. Окремі компоненти електронотранспортного ланцюга діють як протонні помпи, що спричиняють векторний (перпендикулярний площині мембрани) транспорт протонів, спрямований у напрямку "матрикс → зовнішня поверхня мембрани" Спроможність мітохондріальних переносників електронів до транслокації протонів через мембрану зумовлюється особливостями їх внутрішньомембранної топографії. Вважають, що дихальний ланцюг укладений у спрягаючій мембрані у вигляді трьох окислювально-відновлювальних "петель", що відповідають трьом комплексам переносу електронів - І, III та IV і транспортують два іони Н+ з матриксу в зовнішнє середовище.
3. Електрохімічний потенціал протонів на спрягаючих мембранах, який створюється завдяки дії протонних помп дихального ланцюга, є рушійною силою синтезу АТФ з АДФ та Фн.
4. Існує ферментна система, що використовує енергію електрохімічного протонного потенціалу для синтезу АТФ за рахунок зворотної транслокації протонів через мітохондріальну мембрану в напрямку "зовнішня поверхня → матрикс". Ця ферментна система, яка замикає протонний цикл на спрягаючих мембранах мітохондрій - протонна АТФаза, або АТФ-синтетаза. АТФ-синтетаза є білком з четвертинною структурою, що складається з декількох білкових субодиниць, які утворюють компоненти F0 та F1.
5. Будь-які фізичні, хімічні та біологічні фактори, що пошкоджують цілісність спрягаючих мембран мітохондрій та розсіюють енергію електрохімічного градієнта, порушують синтез АТФ, тобто виступають як роз'єднувачі транспорту електронів та окисного фосфорилування.
Таким чином, згідно з хеміосмотичною теорією, спряження між переносом електронів в дихальному ланцюгу та синтезом АТФ здійснюється за рахунок утворення при функціонуванні протонних помп градієнта концентрації Н+ між двома поверхнями мітохондріальної мембрани. АТФ-синтетаза, транспортуючи протони у зворотному напрямку (за електрохімічним градієнтом) призводить до вивільнення хімічної енергії, за рахунок якої утворюються макроергічні зв'язки АТФ.
36. Інгібітори транспорту електронів та роз’єднувачі окисного фосфорилювання.
Певні хімічні сполуки здатні специфічним чином порушувати електронний транспорт (інгібітори електронного транспорту) та окисне фосфорилування (інгібітори та роз'єднувачі окисного фосфорилування) в мітохондріях. Дані сполуки взаємодіють з певними компонентами дихального ланцюга або системи окисного фосфорилування, порушуючи їх біохімічні функції.
Інгібітори електронного транспорту Сполуки цього класу порушують функціонування дихального ланцюга мітохондрій за рахунок зв'язування з окремими ферментними білками або коферментами, що беруть безпосередню участь у переносі електронів від субстратів біологічного окислення на O2. При надходженні в організм людини або тварин ці речовини діють як клітинні отрути, спричиняючи феномен тканинної гіпоксії.
Ротенон - інгібітор транспорту електронів через НАДН:коензим Q-редуктазний комплекс. Ротенон застосовується як інсектицид.
Амобарбітал (амітал) та близький до нього за структурою секобарбітал (секонал). Ці похідні барбітурової кислоти (барбітурати) застосовуються у фармакології як снодійні засоби. Разом з тим, барбітурати, подібно до ротенону, є активними інгібіторами клітинного дихання, блокуючи електронний транспорт на рівні НАДН:коензим Q-редуктази.
Пієрицидин А - антибіотик, що також блокує НАДН:коензим Q-редуктазний
комплекс за рахунок конкурентної взаємодії з убіхіноном.
Антиміцин А - антибіотик, що блокує дихальний ланцюг мітохондрій на рівні переносу електронів через комплекс III (цитохром b - цитохром с1).
Ціаніди (іони CN-) - потужні клітинні отрути, що є інгібіторами транспорту електронів на термінальній ділянці дихального ланцюга мітохондрій (у цитохромоксидазному комплексі). Іони CN- утворюють комплекси з фери (Fе3+)-формою молекул гему цитохромоксидази, блокуючи їх відновлення до феро (Fе2+)-форм.
Монооксид вуглецю (CO) - інгібує цитохромоксидазу шляхом зв'язування з ділянкою гема, що взаємодіє з молекулою кисню.
Інгібітори окисного фосфорилування
Інгібітори окисного фосфорилування блокують як окислення субстратів, так і фосфорилування АДФ у мітохондріях. Олігоміцин - антибіотик, що протидіє як фосфорилюванню АДФ до АТФ, так і стимуляції поглинання O2, що спостерігається після додавання до мітохондрій АДФ (феномен "дихального контролю"). Механізм дії олігоміцину полягає в інгібуванні функції АТФ-синтетази.
Роз'єднувачі окисного фосфорилування
Сполуки цього класу спричиняють "неконтрольоване" дихання мітохондрій, яке не залежить від функціонування системи фосфорилування АДФ. В присутності роз'єднувачів спостерігається активне поглинання мітохондріями О2, незважаючи на зниження швидкості (або відсутність) генерації АТФ з АДФ та Фн. Згідно з хеміосмотичною теорією, роз'єднувачі спричиняють втрату мембраною протонного потенціалу - рушійної сили генерації макроергічних зв'язків АТФ. До роз'єднувачів окисного фосфорилування належать:
- 2,4-динітрофенол та сполуки, близькі до нього за хімічною структурою (динітрокрезол, пентахлорфенол);
- карбонілціанід-м-хлорфенілгідразон - сполука, що в 100 разів перевищує за специфічною активністю 2,4-динітрофенол.
Здатність роз'єднувати дихання та окисне фосфорилування в мітохондріях мають також гормони щитовидної залози (тироксин, трийодтиронін).
Порушення синтезу АТФ спостерігається в умовах дії на організм людини і тварин багатьох патогенних факторів хімічного (природні та синтетичні токсини), біологічного та фізичного (іонізуюча радіація) походження, які спричиняють роз'єднання дихання та окисного фосфорилування за рахунок порушення спроможності створювати і підтримувати протонний потенціал на спрягаючих мембранах мітохондрій.
37. Мікросомальне окислення: цитохром Р-450; молекулярна організація ланцюга переносу електронів.
У мембранах ендоплазматичного ретикулуму печінки, в мітохондріях і мікросомах кори надниркових, статевих залоз та інших тканин локалізовані ферментні системи, які каталізують монооксигеназні реакції, коли один атом молекули кисню включається в субстрат, а другий – у молекулу води. Оскільки найчастіше субстрат у монооксигеназних реакціях гідроксилюється, цю групу ферментів називають також гідроксилазами. Донором воднів для утворення Н2О замість НАДФН може бути НАДН, ФМНН2, ФАДН2.
Головний компонент монооксигеназ – цитохром Р-450 – названий так тому, що комплекс його відновленої форми з монооксидом вуглецю (II) має максимум поглинання світла при 450 нм. Цитохром Р-450 містить протогем і подібний до цитохромів групи b. Буква “Р” в цитохромі Р-450 походить від американського міста Philadelphia, де він вперше був відкритий. Існує велика кількість ізоформ цитохрому Р-450. Оскільки в процесі один із атомів молекули кисню включається в молекулу води, а другий - в молекулу субстрату, що гідроксилюється, ферментні системи, які каталізують ці реакції, отримали також назву "мікросомальних оксигеназ мішаної функції'' Ферментні системи, що каталізують реакції мікросомального окислення гідрофобних речовин, являють собою електронотранспортні ланцюги, локалізовані в мембранах ендоплазматичного
ретикулуму гепатоцитів та клітин деяких інших органів тварин та людини, що також беруть участь у реакціях детоксикації (кишечник, легені, шкіра, плацента тощо). Компонентами цих ферментних ланцюгів є ФАД-вмісний флавопротеїн, цитохром b5 та кінцева монооксигеназа - цитохром Р-450:
Подібний цитохром Р-450-залежний електронотранспортний ланцюг каталізує реакції окислювального гідроксилування стероїдів (синтезу та біотрансформації), що відбуваються в мітохондріях кори наднирників та статевих залоз.
38. Анаеробне окислення глюкози. Послідовність реакцій та ферменти гліколізу.
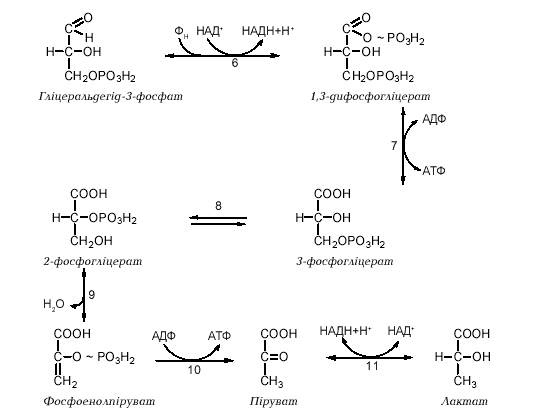
За умов анаеробного гліколізу (наприклад, в інтенсивно працюючих скелетних м`язах або в молочнокислих бактеріях) гліколі-тичний НАДН не віддає свої відновлювальні еквіваленти в дихальний ланцюг мітохондрій, а використовується для відновлення пірувату до L-лактату: Реакція каталізується ферментом лактатдегідрогеназою, яка існує у вигляді п яти різних ізоферментних форм (ЛДГ1—ЛДГ5), що відрізняються за своїми кінетичними властивостями (КМ, Vмах, ступенем алостеричного інгібірування піруватом). Таким чином, ферментативні реакції анаеробного гліколізу майже повністю співпадають із реакціями аеробного гліколізу, відрізняючись лише на етапі, що відбувається після утворення пірувату: при аеробному гліколізі піруват є субстратом перетворення на ацетилкоензим А та подальшого окислення, а при анаеробному гліколізі піруват відновлюється до лактату за рахунок НАДН, що утворився в реакціях гліколітичної оксидоредукції. Інакше кажучи, після утворення пірувату подальше його перетворення може відбуватися за одним із двох альтернативних шляхів, що залежать від стану окислювально-відновлювальних процесів у певній тканині: - в анаеробнихумовах (або в умовах гіпоксії) реокислення гліколітичного НАДН відбувається за рахунок дії лактатдегідрогенази, яка відновлює піруват до лактату; течія лактат-дегідрогеназної реакції в даному напрямку генерує НАД+, що знову використовується для окислення гліцеральдегід-3-фосфату і подальшого накопичення лактату як продукту анаеробного гліколізу. Така послідовністьреакцій найбільш характерна для інтенсивно працюючихскелетних м`язів; крім скелетних м`язів та еритроцитів, клітини деяких інших органів та тканин (головного мозку, шлунково-кишкового тракту, мозкового шару нирок, сітківки та шкіри) частково задовольняють свої енергетичні потреби за рахунок анаеробного гліколізу, утворюючи молочну кислоту.
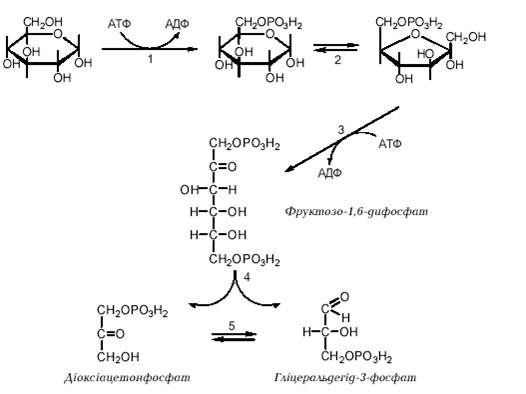
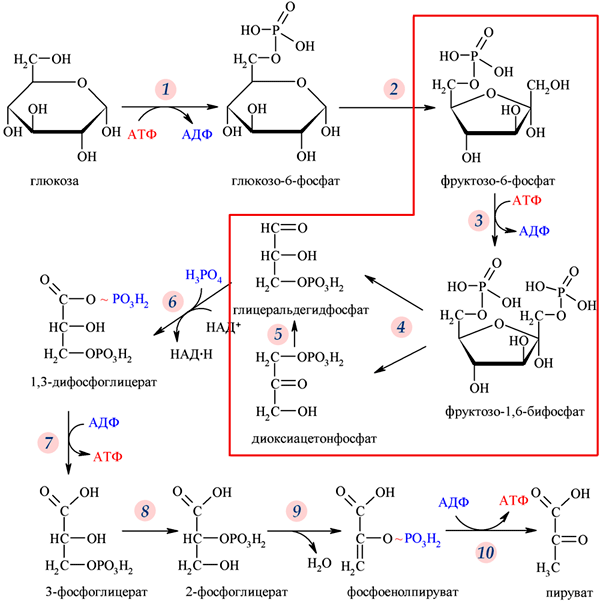
39. Аеробне окислення глюкози. Етапи перетворення глюкози до СО2 і Н2О.
Для більшості тканин людини та вищих тварин в умовах нормальної життєдіяльності характерний аеробний гліколіз, тобто утворення з глюкози пірувату, який у подальшому окислюється до вуглекислого газу й води. Анаеробний гліколіз має місце переважно в м`язах при інтенсивній фізичній діяльності, тобто при відносній кисневій недостатності, та в деяких високоспеціалізованих клітинах (зокрема, в еритроцитах, в яких відсутні мітохондрії) або за певних патологічних умов (клітини злоякісних пухлин). Реакції гліколізу перебігаютьу цитозолі клітини і каталізуються ферментами, що локалізовані в цьому компартменті. Виділяють дві стадії гліколізу: 1. Розщеплення молекули глюкози до двох молекул фосфотріоз (гліцеральдегід-3-фосфату та діоксіацетонфосфату). Ця стадія включає в себе послідовність реакцій, які потребують витрати двох молекул АТФ на кожну молекулу глюкози, що розщеплюється. 2. Перетворення двох молекул фосфотріоз на дві молекули пірувату (або лактату). Ця стадія включає в себе окислювально-відновлювальні реакції ("гліколі-тична оксидоредукція"), які супроводжуються генерацією чотирьох молекул АТР. Таким чином, у результаті розщеплення однієї молекули глюкози в реакціях аеробного або анаеробного гліколізу сумарний вихід АТР складає дві молекули, що можна подати таким рівнянням: D-глюкоза + 2 АДФ + 2 Фн? 2 піруват (лактат) + 2 АТФ
40. Окислювальне декарбоксилювання пірувату. Ферменти, коферменти та послідовність реакцій в мультиферментному комплексі.
Окислювальне декарбоксилювання пірувату каталізується піруватдегідрогеназним комплексом — мультиферментною системою, яка в клітинах еукаріотів міститься в мембранах мітохондрій, а у прокаріотів — у цитоплазмі. До складу цього комплексу входятьтри ферменти, що каталізують три послідовні стадії перетворення пірувату на ацетил-КоА: • піруватдегідрогеназу [КФ 1.2.4.1]; • дигідроліпоїлацетилтрансферазу [КФ 2.3.1.12]; • дигідроліпоїлдегідрогеназу [КФ 1.8.1.4] і п'ять коферментів і простетичних груп: • тіаміндифосфат (ТДФ, або ТПФ); • ліпоєву кислоту(ЛК); • КоА; • ФАД; • НАД+ У результаті цього процесу утворюється ацетилкоензим А — основний субстрат окислення в циклі трикарбонових кислот — та відновлена форма НАД+. Сумарне рівняння окислювального декарбоксилювання пірувату: CH-CO-COOH + НАД+ + КоА-SH -> CH-CO-КоА + НАДН + Н+ + С02 Ферментативні стадії утворення ацетил-КоА з пірувату: Стадія I (декарбоксилювання) — каталізується піруватдегідрогеназою (Е1), коферментом якої є ТДФ. На цій стадії відбувається взаємодія пірувату з С-2 тіазольного кільця молекули тіаміну; в результаті реакції утворюється зв’язаний із ферментом гідроксиетильний похідний тіаміндифосфату: Стадія II — каталізується центральним ферментом комплексу дигідроліпоїлацетилтрансферазою (E2), яка переносить гідроксиетильну групу від ТДФ(Е1) на простетичну групу ферменту E2, що є окисленою формою ліпоєвої кислоти (ЛК); в результаті реакції утворюється ацетилтіоефір відновлених ліпоїльних груп ферменту E2, що містить макроергічний зв’язок: Стадія ІІІ — дигідроліпоїлацетилтрансфераза переносить ацетильну групу від відновленої ліпоєвої кислоти на коензим А: Стадія IV — окислення відновленої форми ферменту Е2 ФАД-залежною дигідроліпоїлдегідрогеназою (E3): Стадія V — перенесення атомів водню від відновленої ФАД-групи дигідроліпоїлдегідрогенази на НАД+ з утворенням НАДН: Відновлений НАДН, що утворюється в результаті окислювального декарбоксилювання пірувату, в аеробних умовах окислюється в мітохондріальному електронотранспортному ланцюзі з генерацією шести (2x3) молекул АТФ. Біологічна роль окиснювального декарбоксилювання пірувату полягає: 1) в катаболізмі пірувату до одного з кінцевих продуктів - С02 (виводиться з організму або використовується для синтезу), 2) в утворенні макроергічної сполуки - ацетил-Ко А (піддається подальшому окисненню в циклі трикарбонових кислот або використовується в реакціях анаболізму); 3) в синтезі відновленого еквіваленту - НАДН, який окиснюється в дихальному ланцюгу мітохондрій. Реакції окиснювального декарбоксилювання пірувату регулюються в організмі на рівні піруватдегідрогенази: 1) алостерично (надлишок продуктів реакції ацетил-КоА НАДН інгібує фермент, а проміжний продукт гліколізу фруктозо-1,6-дифосфат, НАД+, КоА - є активаторами піруватдегідрогенази); 2) шляхом хімічної модифікації - фосфорилування. Фосфорильована форма піруватдегідрогенази є неактивною, а дефосфорильована — активна. Тому інсулін підвищує активність комплексу, а глюкагон, адреналін (діють за аденілатциклазним каскадом) гальмують активність ферменту. Окислювальне декарбоксилювання пірувату – єдиний шлях його катаболізму, тому недостатність вітамінів (у перше чергу вітаміну В1) призводить до порушення процесу, зниження утворення АТФ і проявляється порушенням діяльності ЦНС. Піруватдегідрогеназна недостатність в організмі призводить до підвищення концентрації лактату, пірувату, аланіну, що супроводжується ацидозом.
в аеробних умовах відбувається окисне декарбоксилювання пірувату доацетил-КоА, який у подальшому окислюється до СО, та Н,О в циклі Кребса; НАДН, що утворився при окисленні гліцеральдегід-3-фосфату, віддає свої відновлювальні еквіваленти на дихальний ланцюг мітохондрій через спеціальні човникові механізми; - в анаеробнихумовах (або в умовах гіпоксії) реокислення гліколітичного НАДН відбувається за рахунок дії лактатдегідрогенази, яка відновлює піруват до лактату; течія лактат-дегідрогеназної реакції в даному напрямку генерує НАД+, що знову використовується для окислення гліцеральдегід-3-фосфату і подальшого накопичення лактату як продукту анаеробного гліколізу. Така послідовністьреакцій найбільш характерна для інтенсивно працюючихскелетних м'язів; крім скелетних м'язів та еритроцитів, клітини деяких інших органів та тканин (головного мозку, шлунково-кишкового тракту, мозкового шару нирок, сітківки та шкіри) частково задовольняють свої енергетичні потреби за рахунок анаеробного гліколізу, утворюючи молочну килоту. Човникові механізми. Суть процесу полягає в тому, що НАДН, який утворюється в цитозолі не може самостійно прникнути в мітохондрії для окислення, тому він відновлює у цтиозолі певну речовину, яка йде в мітохондрії і там окислюється (замість НАДН), відновлюючи внутрішньомітохондріальний НАД+, і знову повертається в цитозоль; цикл повторюється. Малат-аспартатна човникова система. У цитозолі відновлюється оксалоацетат до малату. Малат іде в мітохондрії і там окислюється знову до оксалоацетату, відновлюючи НАД+. Гліцерофосфат на човникова система. У цитозолі відновлюється діоксиацетонфосфат до гіцерол-3-ф, який іде в мітохондрії і там окислюється до діоксиацетонфосфату.
41. Порівняльна характеристика біоенергетики аеробного та анаеробного окислення глюкози, ефект Пастера.
У процесі анаеробного окислення глюкози витрачається 2 молекули АТФ (при фосфорилюванні глюкози з утворенням глюкозо-6-фосфату і при перетворенні фруктозо-6-фосфату на фруктозо-1,6-дифосфат), а синтезується 4 АТФ (по дві у реакціях гліцеральдегід-3-фосфат → 3-фосфогліцерат та фосфоенолпіруват → піруват). Різниця у затраченій і утвореній кількості АТФ складає +2 молекули. Сумарне рівняння: С6Н12О6 + 2АДФ + 2Фн→ 2С3Н4О3(С3Н6О3) + 2АТФ. При аеробному окисленні глюкози (повне окислення до СО2 і Н2О): генерується 2АТФ на етапі аеробного гліколізу; гліколітичний НАДН за рахунок окислення в мітохондріях дає 2НАДН*3АТФ= 6АТФ; окисне декарбоксилювання ПВК дає 2НАДН, які в мітохондріях окислються з утворенням 2НАДН*3АТФ= 6АТФ; в ЦТК 2 ацетил-КоА (із попереднього декарбоксилування ПВК) дають 2ац-КоА*12АТФ=24АТФ. Сумарне рівняння: С6Н12О6 + 6О2 + 38АДФ + 38Фн→ 6СО2 + 6Н2О + 38АТФ. Ефект Пастера – не утворення лактату в присутності О2, через те, що в умовах активного клітинного дихання пригнічується активність фосфофруктокінази і піруваткінази.
42. Фосфоролітичний шлях розщеплення глікогену в печінці та м'язах. Регуляція активності глікогенфосфорилази.
С6Н10О5)n + Н3РО4→ г-1-ф + (С6Н10О5)n-1, Ė глікогенфосфорилаза. Розщеплення розгалужених фрагментів – Ė аміло-1,6-глікозидаза; г-1-ф→ г-6-ф, Ė фосфоглюкомутаза. Утворений г-6-ф у печінці перетворюється на вільну глюкозу (Ė г-6-фосфатаза) і йде в кров, а у м`язах, де немає цього ферменту – використовується для власних потреб клітини. Регуляція: адреналін, глюкагон→ рецептор→ G-білок→ аденілатциклаза (активується)→ синтез цАМФ із АТФ→ цАМФ-залежна протеїнкіназа (активується)→ Кіназа фосфорилази фосфорилюється за допомогою АТФ (активується)→ фосфорилаза b (глікогенфосфорилаза) фосфорилюється за допомогою АТФ (активується) – перетвор. на фосфорилазу а→ глікоген+Фн→г-1-ф→г-6-ф→глюкоза.
Гликогенфосфорилаза (glycogen phosphorylase) - Фермент, катализирующий расщепление гликогена до глюкозы, что связано с процессами сокращения мышц (мышечная гликогенфосфорилаза), контролем уровня глюкозы в крови(печеночная гликогенфосфорилаза), межклеточной передачи энергии в условиях аноксии (гликогенфосфорилаза мозга). Соответственно, у млекопитающих гликогенфосфорилаза представлена тремя изоферментами, кодируемыми у ряда видов несцепленными генами (например, у мыши гены гликогенфосфорилазы локализованы на хромосомах 19, 12 и 2). Каждый из изоферментов представлен гомодимером (молекулярная масса около 100 кД), могут образовывать гетеродимеры друг с другом.
43. Біосинтез глікогену: ферментативні реакції, фізіологічне значення. Регуляція активності глікогенсинтази.
Глюкоза→ г-6-ф, Ė гексокіназа або глюкокіназа; г-6-ф→ г-1-ф, Ė фосфоглюкомутаза. Утворюється активна форма глюкози, для синтезу глікогену: г-1-ф + УТФ↔ УДФ-1-г, Ė УДФ-глюкозопірофосфорилаза. Синтез глікогену (утворення α-1,4-глікозидних зв`язків): УДФ-1-г + термінальний залишок глюкози (С6Н10О5)n→ УДФ + (С6Н10О5)n+1; Ė глікогенсинтаза (УДФ-глікогентрансфераза). Розгалуження утворюються переносом кінцевого фрагменту полісахариду з 6-7 мономерів на кілька мономерів далі; Ė аміло(1,4-1,6)глікозилаза. Регуляція: (див. «регуляція» 62). У той час, як глікогенфосфорилаза фосфорилюється і стає активною – глікогенсинтаза теж фосфорилюється (цАМФ-залежною протеїнкіназою), але в цьому стані – неактивна. У гепатоцитах глікоген зберігається у вигляді великих цитоплазматичних гранул. Елементарна так звана β-частинка, що є однією молекулою глікогену, має діаметр близько 21 нм і включає до 55 000 залишків глюкози та має 2000 нередукуючих кінців. 20-40 таких часточок разом утворюють α-розетки, які можна бачити у під мікроскопом у тканинах тварин, яких добре годують. Проте вони зникають після 24-годинного голодування. Глікогенові гранули – це складні агрегати, до складу яких крім самого глікогену входять ферменти, що синтезують і розщеплюють його, а також регуляторні молекули (рис. 3).
Глікоген у м’язах слугує джерелом швидкої енергії як за аеробного, так і за анаеробного метаболізму. Його запаси можуть бути вичерпані за одну годину інтенсивного фізичного навантаження
44. Механізми реципрокної регуляції глікогенолізу та глікогенезу за рахунок каскадного цАМФ-залежного фосфорилювання ферментних білків.
Субстратом гліколізу в м'язах служать глюкоза, яка надходить із крові, і глюкозні залишки депонованого глікогену. Внаслідок послідовної дії глікогенфосфорилази і фосфоглюкомутази глюкозні залишки глікогену перетворюються в глюкозо-6-фосфат, який далі включається в процес гліколізу:
За умов глікогенолізу АТФ затрачається тільки один раз для утворення фруктозо-1,6-дифосфату. Якщо ж врахувати затрати АТФ для біосинтезу глікогену (дві молекули АТФ для включення одного залишку глюкози), тоді чистий вихід складає тільки 1 молекулу АТФ на 1 залишок глюкози. Витрачання АТФ для синтезу глікогену в м'язах має місце в стані спокою, коли депонування глікогену достатньо забезпечене киснем і енергією. А під час інтенсивного фізичного навантаження анаеробний розпад глікогену до молочної кислоти зумовлює більший вихід АТФ, ніж розпад глюкози. Глікогене́з — процес біосинтезу глікогену, що відбувається у клітинах тих живих організмів, які використовують цей полісахарид для зберігання глюкози (тварин, грибів та багатьох прокаріот). У ссавців протікає з різною інтенсивністю у всіх тканинах, але найбільш виражено у печінці та скелетних м'язах. Субстратом для глікогенезу єуридиндифосфатглюкоза, тобто він здійснюється шляхом відмінним від деградації глікогену — глікогенолізу, основним продуктом якого є глюкозо-1-фосфат. Ключовимферментом глікогенезу є глікогенсинтаза.
45. Роль адреналіну, глюкагону та інсуліну в гормональній регуляції обміну глікогену в м'язах та печінці.
Адреналін стимулює розпад і гальмує синтез глікогену в печінці, скелетних м'язах, міокарді. Секреція його у стресових ситуаціях зумовлює вивільнення глюкози із печінки в кров для постачання інших органів, а в м'язах – розпад глікогену до молочної кислоти з виділенням енергії, що забезпечує швидке зростання м'язової активності. Глюкагон стимулює розпад глікогену печінки, але не впливає на глікоген м'язів. Секретується підшлунковою залозою при зниженні концентрації глюкози в крові.
Гормон підшлункової залози інсулін стимулює надходження глюкози в клітини і синтез глікогену. Механізми його дії ще до кінця не з'ясовані. Одним із них є активація інсуліном фосфодіестерази цАМФ, що приводить до зниження внутрішньоклітинного рівня цАМФ, у результаті стимулюється утворення неактивної фосфорилази й активної глікогенсинтази. У гепатоцитах інсулін підвищує активність глюкокінази.
46. Генетичні порушення метаболізму глікогену (глікогенози, аглікогенози).
Глікогенози – спадкові хвороби, молекулярною основою виникнення яких є уроджена недостатність синтезу певних ферментів глікогенолізу, пов’язана з дефектами в генетичній системі клітин. При глікогенозах у внутрішніх органах та тканинах (здебільшого в печінці, м’язах, клітинах крові) спостерігається накопичення аномально надмірної кількості глікогену, іноді зі зміненою молекулярною структурою, який не може використовуватися у метаболічних процесах. Клінічно глікогенози проявляються важкою гіпоглюкоземією внаслідок нездатності глікогену печінки розщеплюватися з вивільненням молекул глюкози.
Аглікогенози – спадкові хвороби накопичення глікогену, молекулярною основою яких є генетичні дефекти, що призводять до порушення утворення ферменту глікогенсинтази.
47. Глюконеогенез: субстрати, ферменти та фізіологічне значення процесу.
Глюконеогенез – це процес синтезу глюкози з невуглеводних субстратів. Такими попередниками глюкози є лактат, піруват, більшість амінокислот, гліцерин, проміжні продукти циклу лимонної кислоти. Відбувається глюконеогенез у печінці й, невеликою мірою, в кірковій речовині нирок. Завдяки цьому процесу підтримується концентрація глюкози в крові після того, як вичерпаються запаси глікогену при вуглеводному чи повному голодуванні. Надзвичайно важливе значення глюконеогенезу для організму тварин і людини зумовлюється тим, що мозок має дуже малі запаси глікогену і глюкоза крові служить основним джерелом енергії для нього. При зменшенні концентрації глюкози в крові нижче певної критичної межі порушується функціонування мозку і може настати смерть. Механізм глюконеогенезу також забезпечує видалення з крові таких продуктів тканинного метаболізму, як лактат і гліцерин.У клітинах печінки здійснюється координована регуляція гліколізу і глюконеогенезу відповідно до фізіологічних потреб усього організму. Система контролю включає субстрати і проміжні продукти процесів, регуляторні ферменти та їх ефектори, гормони. Підкреслимо, що регуляторними ферментами глюконеогенезу і гліколізу є ті, що каталізують незворотні реакції і не беруть участі у протилежному процесі. Наприклад, піруваткарбоксилаза (активатор – ацетил-КоА) і фруктозодифосфатаза (активатор – цитрат, інгібітори – АМФ і фруктозо-2,6-дифосфат). На рис. показані регуляторні пункти гліколізу і глюконеогенезу та контрольні чинники.
Деякі ефектори одночасно впливають на активність регуляторних ферментів в обох процесах. Так, ацетил-КоА служить активатором піруваткарбоксилази і інгібітором піруваткінази. Крім того, ацетил-КоА є інгібітором піруватдегідрогеназного комплексу і, сповільнюючи розпад пірувату до ацетил-КоА, сприяє переходу пірувату в глюкозу. АМФ інгібує фруктозо-дифосфатазу і разом з тим активує відповідний фермент гліколізу – фосфофруктокіназу. Навпаки, цитрат – активатор фосфатази й інгібітор кінази..Глюконеогенез стимулюється не тільки активаторами піруваткарбоксилази і фруктозодифосфатази, а й інгібіторами регуляторних ферментів гліколізу (АТФ, аланін, жирні кислоти). Таким чином, коли в клітині є достатня концентрація палива для циклу лимонної кислоти (ацетил-КоА, оксалоацетату, цитрату, жирних кислот, аланіну) чи висока концентрація АТФ і низькі –АДФ та АМФ, посилюється біосинтез глюкози і гальмується гліколіз. У протилежній ситуації стимулюється гліколіз і гальмується глюконеогенез.. На глюконеогенез впливають такі гормони, як глюкагон й інсулін підшлункової залози та глюкокортикоїди кори надниркових залоз. При голодуванні глюкагон посилює розпад жирів у жировій тканині. Жирні кислоти надходять у печінку, де розпадаються до ацетил-КоА. Швидкість окиснення ацетил-КоА в циклі лимонної кислоти відстає від швидкості його утворення, і підвищений рівень ацетил-КоА в клітині активує піруваткарбоксилазу. В результаті посилюється глюконеогенез.
48. Глюкозо-лактатний (цикл Корі) та глюкозо-аланіновий цикли.
важливим субстратом глюконеогенезу є молочна кислота, яка накопичується в організмі під час інтенсивної м'язової роботи внаслідок анаеробного розпаду глікогену. У період відновлення після напруженої роботи молочна кислота переноситься кров'ю з м'язів до печінки, де під дією лактатдегідрогенази окиснюється до пірувату. Частина останнього використовується для глюконеогенезу, а частина розпадається аеробним шляхом, забезпечуючи процес глюконеогенезу АТФ. Глюкоза потрапляє назад у скелетні м'язи і застосовується для відновлення запасу глікогену. Поєднання процесу анаеробного гліколізу в скелетних м'язах і глюконеогенезу в печінці називається циклом Корі.
studopedia.su - Студопедия (2013 - 2025) год. Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав!Последнее добавление
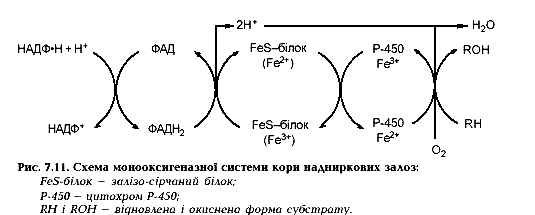 У мембранах ендоплазматичного ретикулуму печінки, в мітохондріях і мікросомах кори надниркових, статевих залоз та інших тканин локалізовані ферментні системи, які каталізують монооксигеназні реакції, коли один атом молекули кисню включається в субстрат, а другий – у молекулу води. Оскільки найчастіше субстрат у монооксигеназних реакціях гідроксилюється, цю групу ферментів називають також гідроксилазами. Донором воднів для утворення Н2О замість НАДФН може бути НАДН, ФМНН2, ФАДН2.
Головний компонент монооксигеназ – цитохром Р-450 – названий так тому, що комплекс його відновленої форми з монооксидом вуглецю (II) має максимум поглинання світла при 450 нм. Цитохром Р-450 містить протогем і подібний до цитохромів групи b. Буква “Р” в цитохромі Р-450 походить від американського міста Philadelphia, де він вперше був відкритий. Існує велика кількість ізоформ цитохрому Р-450. Оскільки в процесі один із атомів молекули кисню включається в молекулу води, а другий - в молекулу субстрату, що гідроксилюється, ферментні системи, які каталізують ці реакції, отримали також назву "мікросомальних оксигеназ мішаної функції'' Ферментні системи, що каталізують реакції мікросомального окислення гідрофобних речовин, являють собою електронотранспортні ланцюги, локалізовані в мембранах ендоплазматичного
ретикулуму гепатоцитів та клітин деяких інших органів тварин та людини, що також беруть участь у реакціях детоксикації (кишечник, легені, шкіра, плацента тощо). Компонентами цих ферментних ланцюгів є ФАД-вмісний флавопротеїн, цитохром b5 та кінцева монооксигеназа - цитохром Р-450:
У мембранах ендоплазматичного ретикулуму печінки, в мітохондріях і мікросомах кори надниркових, статевих залоз та інших тканин локалізовані ферментні системи, які каталізують монооксигеназні реакції, коли один атом молекули кисню включається в субстрат, а другий – у молекулу води. Оскільки найчастіше субстрат у монооксигеназних реакціях гідроксилюється, цю групу ферментів називають також гідроксилазами. Донором воднів для утворення Н2О замість НАДФН може бути НАДН, ФМНН2, ФАДН2.
Головний компонент монооксигеназ – цитохром Р-450 – названий так тому, що комплекс його відновленої форми з монооксидом вуглецю (II) має максимум поглинання світла при 450 нм. Цитохром Р-450 містить протогем і подібний до цитохромів групи b. Буква “Р” в цитохромі Р-450 походить від американського міста Philadelphia, де він вперше був відкритий. Існує велика кількість ізоформ цитохрому Р-450. Оскільки в процесі один із атомів молекули кисню включається в молекулу води, а другий - в молекулу субстрату, що гідроксилюється, ферментні системи, які каталізують ці реакції, отримали також назву "мікросомальних оксигеназ мішаної функції'' Ферментні системи, що каталізують реакції мікросомального окислення гідрофобних речовин, являють собою електронотранспортні ланцюги, локалізовані в мембранах ендоплазматичного
ретикулуму гепатоцитів та клітин деяких інших органів тварин та людини, що також беруть участь у реакціях детоксикації (кишечник, легені, шкіра, плацента тощо). Компонентами цих ферментних ланцюгів є ФАД-вмісний флавопротеїн, цитохром b5 та кінцева монооксигеназа - цитохром Р-450: 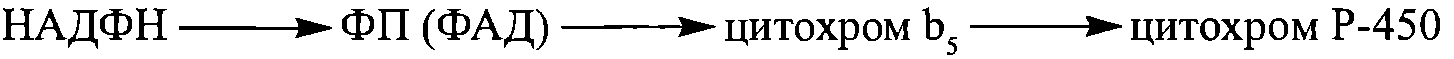 Подібний цитохром Р-450-залежний електронотранспортний ланцюг каталізує реакції окислювального гідроксилування стероїдів (синтезу та біотрансформації), що відбуваються в мітохондріях кори наднирників та статевих залоз.
Подібний цитохром Р-450-залежний електронотранспортний ланцюг каталізує реакції окислювального гідроксилування стероїдів (синтезу та біотрансформації), що відбуваються в мітохондріях кори наднирників та статевих залоз.