Молочна кислота утворюється постійно в еритроцитах, мозковій частині нирок, сітківці ока, а в печінці й корі нирок переходить у глюкозу, яка повинна знову надходити в названі клітини і використовуватись. Таким чином, ця кислота, на відміну від глюкогенних амінокислот, не служить попередником для глюкози крові, що могла б використовуватись у мозку і нервах при голодуванні. Целью глюкозо-аланиновогоцикла также является уборка пирувата, но, кроме этого решается еще одна немаловажная задача – уборка лишнего азота из мышцы. При мышечной работе и в покое в миоците распадаются белки и образуемые аминокислоты рансаминируются с a-кетоглутаратом. Полученный глутамат взаимодействует с пируватом. Образующийся аланин является транспортной формой и пирувата и азота из мышцы в печень. В гепатоците идет обратная реакция трансаминирования, аминогруппа передается на синтез мочевины, пируват используется для синтеза глюкозыГлюкоза, образованная в печени из лактата или аланина, возвращается обратно в мышцы, восстанавливая во время отдыха запасы гликогена.Кроме мышечной работы, глюкозо-аланиновый цикл активируется во время голодания, когда мышечные белки распадаются и многие аминокислоты используются в качестве источника энергии, а их азот необходимо доставить в печень.
49. Глюкоза крові (глюкоземія): нормоглікемія, гіпо- та гіперглікемії, глюкозурія. Цукровий діабет - патологія обміну глюкози.
У нормі через декілька годин після їди концентрація глюкози в крові людини складає 3,33-5,55 ммоль/л. При споживанні вуглеводної їжі вона зростає до 8-9 ммоль/л, а через 2 год повертається до норми. Голодування протягом декількох діб майже не відбивається на рівні глюкози в крові.
Постійність концентрації глюкози дуже важлива з огляду на високу вірогідність порушення функцій головного мозку при гіпоглікемії. Це зумовлюється рядом обставин:
1) енергетичні потреби головного мозку забезпечуються тільки глюкозою (лише на пізній стадії голодування – кетоновими тілами);
2) запаси глікогену в головному мозку дуже незначні;
3) шляхом глюконеогенезу глюкоза в клітинах мозку не синтезується;
4) глюкоза надходить із крові в клітини головного мозку шляхом незалежної від інсуліну дифузії за градієнтом концентрації, при гіпоглікемії надходження стає недостатнім для нормального функціонування мозку. Швидкий розвиток гіперглікемї також може зумовити порушення функцій мозку.
Концентрація глюкози в крові залежить від рівноваги між надходженням її в кров і споживанням тканинами. Оскільки виведення глюкози з організму з сечею в нормі дуже незначне, то підтримка сталості концентрації у відносно вузьких межах за значних коливань надходження з їжею забезпечується процесами обміну в тканинах. Система регуляторних механізмів включає гормони інсулін, глюкагон, адреналін, глюкокортикоїди, а також взаємодії між тканинами (печінкою, м'язами, мозком тощо).
Після споживання вуглеводної їжі підвищена концентрація глюкози в крові стимулює поглинання її тканинами. Швидкість надходження в клітини печінки, м'язів, мозку й інших тканин прямо пропорційна концентрації глюкози в позаклітинній рідині. Крім того, висока концентрація глюкози в циркулюючій крові стимулює секрецію b-клітинами підшлункової залози інсуліну, який підвищує проникність глюкози через клітинні мембрани скелетних м'язів, жирової тканини.
У клітинах інсулін стимулює утилізацію глюкози різними шляхами:
А. У печінці й м'язах синтезується глікоген (інсулін індукує синтез глюкокінази печінки, активує гексокіназу і глікогенсинтазу).
Б. У жировій тканині й печінці глюкоза перетворюється в жирні кислоти, які утворюють тканинні резерви у вигляді тригліцеридів жирової клітковини.
В. Для всіх органів і тканин у період травлення й абсорбції катаболізм глюкози служить основним джерелом енергії. Посилюються гліколіз і аеробний розпад глюкози до СО2 і Н2О. Так, після прийому їжі наближення дихального коефіцієнта до одиниці вказує на більшу інтенсивність окиснення глюкози. Величина катаболізму вуглеводів буде залежати від потреби організму в енергії. Крім того, в цей період високе відношення інсулін/глюкагон в крові стримує глюконеогенез. У результаті концентрація глюкози в крові наближається до норми, опускаючись іноді нижче вихідного рівня. Секреція інсуліну поступово припиняється.
При припиненні надходження вуглеводів їжі концентрація глюкози в крові протягом декількох днів майже не знижується завдяки двом процесам: розпаду глікогену печінки і глюконеогенезу. Зменшення концентрації глюкози в крові до нижньої межі норми ініціює виділення підшлунковою залозою глюкагону, який активує фосфорилазу печінки. Зростають розпад глікогену і вихід глюкози в кров. Розпад глікогену печінки підтримує нормальний рівень глюкози в крові не більше 24 год, але вже через 5-6 год після прийому їжі починається повільне підвищення глюконеогенезу із амінокислот і гліцерину, а через 24 год глюконеогенез перебігає з максимальною активністю. Разом із глюкагоном, який активує ферменти глюконеогенезу, включаються глюкокортикоїди, які стимулюють синтез ферментів глюконеогенезу в печінці й посилюють розпад білків у інших тканинах, що забезпечує процес глюконеогенезу субстратами. Внаслідок низького відношення в крові інсулін/глюкагон під час голодування глюкоза не захоплюється печінкою, скелетними м'язами, міокардом, жировою тканиною. Перераховані чинники забезпечують в умовах голодування надходження глюкози в головний мозок у необхідній кількості. При тривалому голодуванні головний мозок, як і інші тканини, використовує як джерело енергії кетонові тіла.
Крім глюкагону і глюкокортикоїдів, концентрацію глюкози в крові підвищує ще ряд гормонів. Адреналін – гормон мозкової частини надниркових залоз – виділяється в стресових ситуаціях і через каскадний механізм викликає швидкий і сильний розпад глікогену печінки до вільної глюкози. Підвищенням рівня глюкози в крові супроводжується дія гормону росту, адренокортикотропіну, тироксину. Таким чином, концентрацію глюкози в крові знижує тільки інсулін, а підвищує ряд гормонів. Існування групи надійних дублюючих механізмів підкреслює той факт, що найближчі результати гіпоглікемії небезпечніші, ніж наслідки гіперглікемії.
Узгоджена дія різних гормонів зумовлює досконалість регуляції гомеостазу глюкози, забезпечує пристосування обміну вуглеводів у всьому організмі до змін харчування, фізичної активності й інших фізіологічних умов.
Підвищена концентрація глюкози в крові внаслідок споживання вуглеводної їжі (аліментарна гіперглікемія) і внаслідок стресу (емоційна гіперглікемія) швидко знижується до норми. Стійка гіперглікемія може розвинутись при цукровому діабеті, який виникає в результаті абсолютної чи відносної недостатності інсуліну. Інші причини гіперглікемії – надлишкова секреція гормону росту, глюкокортикоїдів, іноді ураження ЦНС, порушення мозкового кровообігу, захворювання печінки, підшлункової залози.
Гіперглікемію при цукровому діабеті можна розглядати як корисне пристосування, яке сприяє використанню глюкози клітинами мозку, міокарда, еритроцитами, тобто інсулінонезалежними тканинами. Однак у скелетні м'язи, печінку та інші інсулінозалежні тканини глюкоза не надходить. При високій концентрації глюкози в крові підвищується швидкість зв'язування її з білками (глікозилювання білків), що зумовлює порушення їх функцій, тому тривала гіперглікемія викликає ряд віддалених ускладнень цукрового діабету
При діагностиці цукрового діабету кров для аналізу краще брати після голодування протягом хоча б 10 год. Концентрація глюкози в плазмі крові, взятої натщесерце, вища за 8 ммоль/л, свідчить про вірогідність цукрового діабету. Якщо концентрація глюкози знаходиться в межах 6‑8 ммоль/л, то досліджують кров після цукрового навантаження (дають випити 75 г глюкози, розчиненої у воді). Концентрація через 2 год після навантаження 10 ммоль/л і вище вказує на цукровий діабет, а концентрація від 8 до 10 ммоль/л – на знижену толерантність до глюкози. У частини осіб із порушеною толерантністю до глюкози можливий розвиток діабету.
50. Гормональна регуляція концентрації та обміну глюкози крові.
У нормі через декілька годин після їди концентрація глюкози в крові людини складає 3,33-5,55 ммоль/л. При споживанні вуглеводної їжі вона зростає до 8-9 ммоль/л, а через 2 год повертається до норми. Голодування протягом декількох діб майже не відбивається на рівні глюкози в крові.
Постійність концентрації глюкози дуже важлива з огляду на високу вірогідність порушення функцій головного мозку при гіпоглікемії. Це зумовлюється рядом обставин:
1) енергетичні потреби головного мозку забезпечуються тільки глюкозою (лише на пізній стадії голодування – кетоновими тілами);
2) запаси глікогену в головному мозку дуже незначні;
3) шляхом глюконеогенезу глюкоза в клітинах мозку не синтезується;
4) глюкоза надходить із крові в клітини головного мозку шляхом незалежної від інсуліну дифузії за градієнтом концентрації, при гіпоглікемії надходження стає недостатнім для нормального функціонування мозку. Швидкий розвиток гіперглікемї також може зумовити порушення функцій мозку.
Концентрація глюкози в крові залежить від рівноваги між надходженням її в кров і споживанням тканинами. Оскільки виведення глюкози з організму з сечею в нормі дуже незначне, то підтримка сталості концентрації у відносно вузьких межах за значних коливань надходження з їжею забезпечується процесами обміну в тканинах. Система регуляторних механізмів включає гормони інсулін, глюкагон, адреналін, глюкокортикоїди, а також взаємодії між тканинами (печінкою, м'язами, мозком тощо).
51. Пентозофосфатний шлях окислення глюкози: схема процесу та біологічне значення.
Основний шлях катаболізму глюкози в організмі людини і тварин – це поєднання гліколізу і циклу лимонної кислоти. Проте існують другорядні шляхи, які виконують специфічні функції. Один із них (пентозофосфатний) включає перетворення глюкозо-6-фосфату в пентозофосфати і СО2 і таким чином забезпечує клітини рибозо-5-фосфатом для синтезу нуклеотидів і нуклеїнових кислот. Крім того, ПФШ постачає відновлену форму НАДФН, необхідну для реакцій відновлення під час синтезу жирних кислот і стероїдних сполук, для мікросомального окиснення. ПФШ складається з окисної і неокисної стадій. Усі реакції відбуваються в цитоплазмі клітин.
У першій реакції окисної стадії глюкозо-6-фосфат окиснюється НАДФ+-залежною глюкозо-6-фосфатдегідрогеназою з утворенням НАДФН і 6-фосфоглюконолактону. Останній гідролізується лактоназою до 6-фосфоглюконової кислоти. Тому ПФШ називають також фосфоглюконатним. Далі фосфоглюконатдегідрогеназа каталізує одночасне дегідрування і декарбоксилювання 6-фосфоглюконату. Перший атом вуглецю гексозофосфату звільняється у вигляді СО2. Утворюються пентоза (рибулозо-5-фосфат) і друга молекула НАДФ. Ці три реакції окисної стадії незворотні.
Неокисна стадія ПФШ включає зворотні реакції ізомеризації пентоз і переходу їх у фруктозо-6-фосфат. Перетворення здійснюються через проміжні сполуки – гептозу (седогептулозо-7-фосфат), тетрозу (еритрозо-4-фосфат), тріозу (гліцеральдегід-3-фосфат) – під дією ферментів транскетолази і трансальдолази, які каталізують реакції перенесення, відповідно дво- і тривуглецевого фрагмента з одного вуглеводу на інший. Коферментом транскетолази є тіаміндифосфат. Утворений фруктозо-6-фосфат перетворюється в глюкозо-6-фосфат або розпадається шляхом гліколізу. Таким чином, ПФШ може переходити в гліколітичний. Так здійснюється утилізація пентоз, утворених в окисній стадії ПФШ, і рибозо-5-фосфату, утвореного при розпаді нуклеїнових кислот.
Реакції неокисної стадії ПФШ перебігають і у зворотньому напрямку. Завдяки цьому пентози можуть синтезуватись із глюкози не через окисну стадію ПФШ, а із проміжних продуктів гліколізу – фруктозо-6-фосфату і гліцеральдегід-3-фосфату. Сумарний перехід п'яти молекул фруктозо-6-фосфату дасть шість молекул рибозо-5-фосфату. Такий напрямок реакцій неокисної стадії ПФШ має місце в тканинах, в яких потреба в пентозах більша, ніж потреба в НАДФН.
На відміну від гліколізу і циклу лимонної лимонної кислоти, ПФШ може функціонувати в різних тканинах в декількох варіантах. Вибір напрямку і варіанту ПФШ визначається наявністю субстратів і потребою клітини в продуктах.
1. При потребі в НАДФН і рибозо-5-фосфаті ПФШ може закінчуватись утворенням пентози.
2. Коли потреба в НАДФН більша, ніж потреба в пентозах, окисна стадія доповнюється переходом пентоз у глюкозо-6-фосфат, який знову окиснюється до пентоз. Із сумарним процесом краще ознайомитись, розглядаючи 6 молекул глюкозо-6-фосфату.
Звернемо увагу, що всі молекули СО2 утворюються із різних молекул глюкозо-6-фосфату.
Значною потребою в НАДФН відрізняються жирова тканина, молочна залоза в період лактації, печінка (для синтезу жирних кислот), кора надниркових залоз і сім'яники (для синтезу стероїдних гормонів), еритроцити (для відновлення окисненого глутатіону). Тому в цих тканинах велика частина глюкози окиснюється пентозофосфатним шляхом.
3. Коли потреба в рибозо-5-фосфаті для синтезу нуклеотидів значно більша, ніж потреба в НАДФН, то відбувається його утворення з фруктозо-6-фосфату реакціями неокисної стадії ПФШ. Величини відносних вкладів окисного і неокисного шляхів утворення пентоз у тканинах організму вивчені недостатньо.
4. Рибозо-5-фосфат, що виникає при розпаді нуклеотидів і нуклеїнових кислот, через реакції неокисної стадії ПФШ включається в гліколіз та аеробний розпад і, таким чином, може служити джерелом енергії.
Регуляторний фермент ПФШ – глюкозо-6-фосфатдегідрогеназа. Активність його гальмується продуктом реакції – НАДФН. Таким чином, інтенсивність ПФШ залежить від швидкості використання НАДФН у реакціях анаболізму і контролюється відношенням у клітині НАДФ+/НАДФН.
Для глюкозо-6-фосфатдегідрогенази еритроцитів відкрита велика кількість варіантів (приблизно 80), які відрізняються за каталітичною активністю, спорідненістю із субстратами, чутливістю до температури, рН, інгібіторів, електрофоретичною рухомістю. Активність ферменту може бути зниженою різною мірою, аж до нульової. Наявність у людини варіанту глюкозо-6-фосфатдегідрогенази з активністю, близькою до нуля, проявляється клінічно гемолітичною анемією. При вираженій недостатності ферменту (активність нижча 10 %) анемія розвивається лише під впливом деяких лікарських середників (протималярійних, сульфаніламідних та ін.). У людей із менш вираженою недостатністю ферменту клінічні прояви, як правило, відсутні. Механізм розвитку гемолітичної анемії при цій спадковій патології полягає в наступному. В еритроцитах НАДФ переводить окиснений глутатіон у відновлений. Останній попереджує пероксидне окиснення ліпідів мембран, захищає від окиснення SH-групи білків, підтримує відновлений стан заліза в гемоглобіні. Знижена продукція НАДФН внаслідок вираженої недостатності глюкозо-6-фосфатдегідрогенази зумовлює зниження рівня в еритроцитах відновленого глутатіону і, як наслідок, гемоліз еритроцитів.
52. Метаболічні шляхи перетворення фруктози та галактози; спадкові ензимопатії їх обміну.
Шляхом гліколізу розщеплюється не тільки глюкоза, а й інші моносахариди – фруктоза, галактоза, маноза. У першій реакції моносахариди фосфорилюються через взаємодію з АТФ, а далі по-різному перетворюються в проміжні продукти гліколізу.
Фосфорилювання фруктози каталізують два ферменти – гексокіназа і фруктокіназа. Неспецифічна гексокіназа, яка діє на більшість гексоз, перетворює фруктозу у фруктозо-6-фосфат:
Гексокіназа має низьку спорідненість із фруктозою, тому цим шляхом фосфорилюється незначна її кількість.
Фермент печінки фруктокіназа каталізує фосфорилювання фруктози за першим атомом вуглецю:
Фруктозо-1-фосфат, на відміну від фруктозо-6-фосфату, не перетворюється у фруктозо-1,6-дифосфат, а відразу розщеплюється на 2 тріози – діоксіацетонфосфат і гліцеральдегід. Реакцію каталізує альдолаза фруктозо-1-фосфату. Гліцеральдегід фосфорилюється при взаємодії з АТФ під дією тріозокінази і таким чином обидві тріози переходять на шлях гліколізу. Оскільки перетворення фруктози в тріози здійснюється без участі двох регуляторних реакцій гліколізу (гексокіназної і фосфофруктокіназної), які обмежують швидкість процесу, катаболізм фруктози відбувається значно швидше, ніж глюкози.
Відомі два спадкові порушення обміну фруктози – есенціальна фруктозурія і непереносимість фруктози (рис.).
У першому випадку зростає її вміст у крові й сечі. Така фруктозурія не супроводжується патологічними ознаками і важливого клінічного значення не має. У другому випадку генетичний дефект альдолази фруктозо-1-фосфату зумовлює суттєві порушення обміну вуглеводів, гіпоглікемію, ураження печінки. При споживанні фруктози накопичується фруктозо-1-фосфат, який гальмує активність глікогенфосфорилази, ряду ферментів гліколізу і глюконеогенезу. Хвороба звичайно виявляється під час переходу від грудного годування дітей до їжі, що містить цукор, коли спостерігаються напади блювання і судоми після їди. При непереносимості фруктози необхідно різко обмежити її споживання.. При спадковій галактоземії відсутня галактозо-1-фосфат-уридилтрансфераза, що зумовлює накопичення в крові й тканинах галактозо-1-фосфату, вільної галактози і спирту дульциту – продукту відновлення галактози. Високий їх вміст діє токсично, в немовлят після споживання молока відзначаються блювання і пронос. Швидко збільшується печінка, погіршується зір із-за катаракти, затримується розумовий розвиток. Раннє виявлення захворювання і вилучення галактози (лактози молока) з їжі дозволяють не допустити появи клінічних симптомів і забезпечити нормальний розвиток дитини.
Реакції катаболізму триацилгліцеролів
Ферментативний гідроліз (ліполіз) триацилгліцеролів в адипоцитах та інших кліти-
нах, де накопичуються нейтральні жири, є фізіологічним механізмом, шо має суттєве
значення як постачальник хімічної енергії, особливо в умовах вичерпання вуглевод-
них резервів та при стресових ситуаціях. Процес розщеплення триацилгліцеролів із
вивільненням жирних кислот, які виходять у кров, отримав назву мобілізації жирних
кислот із жирової тканини.
Внутрішньоклітинний ліполіз триащиюііцеролів (ГГ) здійснюється в декілька стадій,
продуктами яких є діацилгліцероли (дигліцериди - ДГ), моноаішлгліцероли (монопліце-
риди - МГ), гліцерол та вільні жирні кислоти:
Вільні жирні кислоти (неетерифіковані жирні кислоти - НЕЖК) с субстратами
окислення для клітин багатьох тканин, зокрема міокарда, скелетної, гладенької мус-
кулатури тощо, крім головного мозку. Після надходження в плазму крові нерозчинні у
водній фазі плазми крові високомолекулярні жирні кислоти транспортуються у зв’язаній із сироватковим альбуміном молекулярній формі.
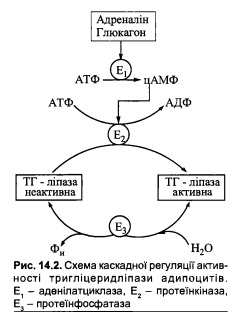
54. Нейрогуморальна регуляція ліполізу за участю адреналіну, норадреналіну, глюкагону та інсуліну).
Регуляція метаболізму ліпідів здійснюється за рахунок впливу фізіологічних та біо-
хімічних ефектів нейроендокринної системи відносно швидкість-лімітуючих реакцій
обміну даного класу біомолекул. Певні гормони та нейромедіатори впливають на ка-
талітичну активність і внутрішньоклітинну концентрацію ключових ферментів розщеплення й біосинтезу ліпідів. Найбільш вивченою є нейрогуморальна регуляція внутрішньоклітинного гідролізу або ліполізу резервних триацилгліцеролів.
Адреналін, норадреналін, глюкагон
Адреналін та норадреналін - катехоламіни, що активують ліполіз у жировій тка-
нині за рахунок стимуляції цАМФ-залежного каскадного механізму регуляції актив-
ності ТГ-ліпази адипоцитів. Ліполітична дія цих гормонів реалізується за умов фізіо-
логічних (фізичне напруження, зниження температури навколишнього середовища)
та психологічних (страх, тривога) стресів, що супроводжуються секрецією з мозково-
го шару надниркових залоз адреналіну, а також стимуляцією симпатичної нервової
системи та вивільненням у синапсах нейронів норадреналіну, які взаємодіють із ад-
ренергічними рецепторами мембран адипоцитів.
Глюкагон - панкреатичний гормон, що стимулює ліполітичну систему в жировій
тканині за механізмом, подібним до дії катехоламінів, тобто за рахунок підвищення в
адипоцитах вмісту цАМФ, пов’язаного з активацією аденілатциклази. Дія глюкагону
проявляється в умовах зниження концентрації глюкози в крові через зменшення її над-
ходження з кишечнику або посиленого використання в тканинах.
У цілому за рахунок розглянутих біохімічних механізмів метаболічні ефекти кате-
холамінів та глюкагону призводять до швидкої стимуляції глікогенолізу в печінці і м’язах та ліполізу в жировій тканині, що забезпечує підвищені енергетичні потреби організму за умов стресу або голодування.
Інсулін
На відміну від зазначених гуморальних факторів, що активують ТГ-ліпазу адипо-
цитів, спричиняючи мобілізацію НЕЖК із жирової тканини, гормон інсулін гальмує
процес ліполізу та вивільнення жирних кислот. Інгібуюча дія інсуліну відносно ліполізу
в адипоцитах реалізується за рахунок двох біохімічних механізмів:
а) зменшення концентрації цАМФ, що може бути пов’язане з активацією фосфо-
дісстсрази цАМФ;
б) збільшення проникності мембран адипоцитів до глюкози, результатом чого с
активація в жировій тканині гліколізу і, відповідно, накопичення гліколітичних мета-
болітів діоксіацстонфосфаіу та 3-фосфогліцсринальдегіду. Ці метаболіти, у свою чергу,
є попередниками гліцсрол-3-фосфату, що необхідний для реетерифікації жирних кис-
лот при біосинтезі триацилгліцеролів. Таким чином, стимульоване інсуліном підвище-
не надходження в адипоцити глюкози переключає метаболізм жирних кислот на вико-
ристання їх здебільшого в синтетичних реакціях і зменшує їх вихід у кров.
Розглянуті біохімічні особливості дії інсуліну пояснюють певні зміни вуглеводного та
ліпідного обмінів, що спостерігаються при голодуванні та цукровану діабеті. Зазначені
стани, для яких властиве зниження концентрації інсуліну в крові, характеризуються також зниженням надходження в адипоцити глюкози, що, зменшуючи шюкозозалежне гальмування мобілізації жирних кислот (див. вище), сприяє їх виходу в плазму крові та використанню іншими тканинами як енергетичного джерела. Введення глюкози та інсуліну хворим на цукровий діабет або експериментальним тваринам, що голодують, спричиняє інгібування ліполізу в адипоцитах і протидіє надмірному збільшенню НЕЖК у плазмі крові.
Соматотропін - гормон передньої частки гіпофіза, який також стимулює ліполіз
у жировій тканині за умов голодування, але його ліполітична дія суттєво відрізняєть-
ся від дії катехоламінів та глюкагону. Соматотропін спричиняє підвищення процесів
ліполізу за рахунок підвищення синтезу відповідних ферментних білків. Метаболічні
ефекти соматотропіну розвиваються повільно, що свідчить про його значення в по-
ступовій адаптації до голодування.
Ліполіз в інших тканинах (м’язах, печінці) регулюється за подібними нейрогу-
моральними механізмами.
55. Реакції окислення жирних кислот (β-окислення); роль карнітину в транспорті жирних кислот в мітохондрії.
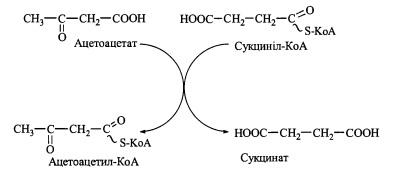
57. Кетонові тіла. Реакції біосинтезу та утилізації кетонових тіл, фізіологічне значення.
Утилізація КТ
58. Порушення обміну кетонових тіл за умов патології (цукровий діабет, голодування).
Метаболізм кетонових тіл в умовах патології
У нормі концентрація кетонових тіл у крові та більшості тканин незначна (в се-
редньому 10-20 мг/л). Проте за умов голодування та цукрового діабету створюють-
ся метаболічні умови, за яких кількість кетонових тіл у тканинах різко підвищується
за рахунок значної активації їх синтезу. При цьому значно зростають як концентрація
кетонових тіл у крові (кетонемія), так і їх виділення з сечею (кетонурія).
Біохімічною основою зростання вмісту кетонових тіл в умовах патологи є
зменшення ступеня утилізації ацетил-КоА в циклі трикарбонових кислот
внаслідок порушень вуглеводного обміну.
Справа в тому, шо входження ацетил-КоА в ЦТК залежить від наявності в клітині
достатньої кількості оксалоацстату, необхідного для утворення цитрату. У свою чер-
гу, утворення оксалоацстату, необхідного для нормального функціонування трикарбо-
нового циклу, залежить від кількості пірувату (розділ 10.4), основним постачальником
якого с гліколітичне розщеплення глюкози. В умовах зменшеного надходження в клітину
глюкози (голодування, цукровий діабет) оксалоацетат спрямовується на шлях піюко-
неогенезу і стає недосяжним для взаємодії з ацетил-КоА в цитратсинтазній реакції. У
зазначених метаболічних умовах ацетил-КоА значною мірою використовується для
синтезу кетонових тіл - ацетоацетагу та Р-гідроксибутирагу. Сприяє накопиченню в
клітинах ацетил-КоА також його підвищене утворення при р-окислснні жирних кислот
за рахунок стимуляції в умовах глюкозного голодування ліполізу в жировій тканині. Ці
біохімічні закономірності пояснюють давній вислів "Жири згоряють у полум ї вуг-
леводів”
Введення в організм глюкози (при голодуванні), або глюкози з інсуліном (при цук-
ровому діабеті) підвищує внутрішньоклітинний рівень моносахариду і нормалізує
гліколіз, що призводить до активації утилізації ацетил-КоА в циклі трикарбонових кис-
лот і зменшення утворення кетонових тіл. Проте в умовах відсутності необхідної те-
рапії концентрація ацетоацетату, Р-гідроксибутирату та ацетону в організмі хворих на
цукровий діабет може зростати в десятки разів, супроводжуючись порушенням кис-
лотно-лужного балансу і розвитком метаболічного кетоацидозу, який с небезпечним
для нормального функціонування клітин головного мозку.
59. Біосинтез вищих жирних кислот: реакції біосинтезу насичених жирних кислот (пальмітату) та регуляція процесу.
60. Біосинтез моно- та поліненасичених жирних кислот в організмі людини.
Мононенасичені жирні кислоти
Мононснасичені кислоти - пальмітоолеїнова С16:1, та олеїнова С18:1 містять подвійний зв’язок між 9-м та 10-м атомами вуглецю.
Ці жирні кислоти можуть утворюватися в організмі людини за рахунок дегідрування
відповідних насичених кислот (пальмітинової С16 та стеаринової С18):
C16 - 2H → C16:1
C18 - 2H → C18:1
Утворення зазначеного подвійного зв’язку здійснюється за участю системи десату-рації жирних кислот (ацил-КоА-оксигенази), що належить за механізмом дії до
мікросомальних монооксигеназ (оксигеназ мішаної функції), які потребують для свого функціонування НАДФН (або НАДН) та включають цитохром b5 електроно-транспортного ланцюга, локалізованого в мембранах ендоплазматичного ретикулуму гепатоцитів:
Стеароїл-КоА (С18) + НАДФН + Н+ + O2 Олеїл-КоА (С18,) + НАДФ + 2 Н2O
Поліненасичені жирні кислоти
Поліненасичсні кислоти - лінолева С18:2 та а-ліноленова С18:3 - попередники в утворенні інших, життєво необхідних ацилів, не можуть синтезуватися в клітинах людського організму.
Зазначені ферменти присутні в багатьох рослинних організмах, і тому існує потреба
в постійному надходженні лінолсвої та α-ліноленової кислот в організм як компонентів рослинної їжі, що є незамінними факторами харчування (“есенціальні жирні кислоти”).
У разі надходження цих жирних кислот у складі дієти, ферментні системи ендо-
плазматичного ретикулуму гепатоцитів за розглянутими вище механізмами десату-
рації та елонгації можуть трансформувати лінолеву кислоту в такі поліненасичені кис-
лоти, як γ-ліноленову С18:3 та арахідонову С20:4, а а-ліноленову – в докозангексенову (С22:6) кислоту.
Арахідонова кислота - попередник біологічно активних ейкозаноїдів (простагландинів, простациклінів, тромбоксанів), утворюється з незамінної лінолевої кислоти С18:2 шляхом подовження її вуглецевого ланцюга та утворення додаткових подвійних зв’язків:
C18:2 – 2H +2C – 2H → C20:4
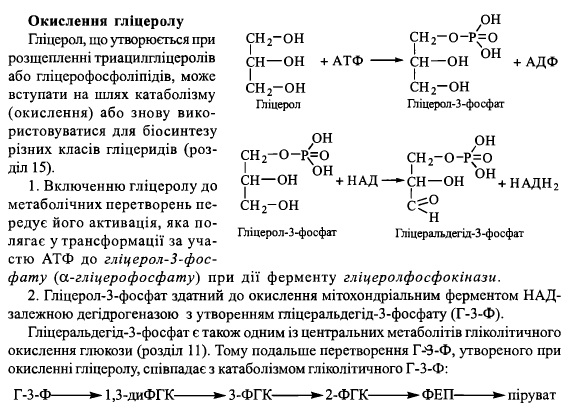
61. Біосинтез триацилгліцеролів та фосфогліцеридів.
Триацилгліцероли (нейтральні жири)- ліпіди, що складають основну частину
харчових ліпідів і в найбільшій кількості представлені в адипоцитах жирової тканини,
де вони виконують функцію резерву метаболічного палива. Кількість нейтральних
жирів в організмі дорослої людини масою 70 кг дорівнює в середньому 10-15 кг. Крім
жирової тканини, біосинтез триацилгліцеролів в обмеженій кількості відбувається та-
кож в інших тканинах, зокрема печінці, кишечнику, молочній залозі в період лактації.
Метаболічними попередниками в біосинтезі триацилгліцеролів є активовані жирні
кислоти (ацил-КоА) та гліцерол-3-фосфат, що, у свою чергу, постачаються за рахунок
окислення глюкози.
Ферментативні реакції синтезу триацилгліцеролів
1. Утворення активованої форми гліцсролу - гліцерол-3-фосфату (а-гліцерофосфату).
studopedia.su - Студопедия (2013 - 2025) год. Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав!Последнее добавление
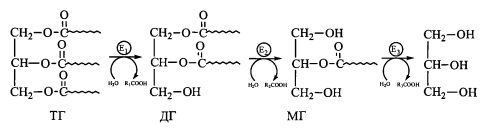 Вільні жирні кислоти (неетерифіковані жирні кислоти - НЕЖК) с субстратами
Вільні жирні кислоти (неетерифіковані жирні кислоти - НЕЖК) с субстратами
 окислення для клітин багатьох тканин, зокрема міокарда, скелетної, гладенької мус-
кулатури тощо, крім головного мозку. Після надходження в плазму крові нерозчинні у
водній фазі плазми крові високомолекулярні жирні кислоти транспортуються у зв’язаній із сироватковим альбуміном молекулярній формі.
окислення для клітин багатьох тканин, зокрема міокарда, скелетної, гладенької мус-
кулатури тощо, крім головного мозку. Після надходження в плазму крові нерозчинні у
водній фазі плазми крові високомолекулярні жирні кислоти транспортуються у зв’язаній із сироватковим альбуміном молекулярній формі.

 Утилізація КТ
Утилізація КТ