Цей процес може відбуватися за одним із двох механізмів:
1.1. Шляхом фосфорилування гліцеролу за участю ферменту гліцеролфосфокінази
1.2. Шляхом відновлення діоксіацетонфосфату - інтермедіату гліколітнчного роз-
іцсшісшія глюкози. Реакція каталізується НАДН-залежною гліцерол-3-фосфат-де-
гідрогеназою (а-гліцерофосфатдегідрогеназою):
Гліцерофосфоліпіди (фосфогліцериди) - фосфатидилхолін, фосфатидил-етаноламін, фосфати дилсерин і кардіол іній - належать до структурних ліпідів, що складають ліпідний матрикс біологічних мембран. Це - складні ліпіди, побудовані на основі гліцеролу, тому перші етапи їх біосинтезу однакові з розглянутими вище фермен-
тативними реакціями утворення триацилгліцеролів, а саме:
Гліцерол → Гліцерол-З-фосфат → Фосфатидна кислота
Після утворення 1,2 діацилптіцеролу реакції синтезу триацилгліцеролів та фосфогліцеридів дивергують.
У разі біосинтезу зазначених фосфогліцеридів до 1,2-діацилгліцеролу, що утворюєть-
ся в результаті гідролізу фосфатидної кислоти, приєднується гідрофільна головка, яка
містить аміноспирт (холін або етаноламін). Особливістю процесу є використання в
реакції активованих форм аміноспиртів - комплексів холіну (етаноламіну) з нуклео-
зиддифосфатом ЦДФ, які утворюються за рахунок таких реакцій:
- активації холіну (етаноламіну) шляхом АТФ-залежного фосфорилування аміно-
спирту;
- взаємодії фосфохоліну (або фосфоетаноламіну) з нуклеозидтрифосфатом ЦТФ з
утворенням ЦДФ-холіну (ЦДФ-етаноламіну);
- При взаємодії ЦДФ-холіну (ЦДФ-етаноламіну) з 1,2-діацилгліцеролом утворю-
ються фосфатидилхолін або фосфатидилетаноламін.
62. Метаболізм сфінголіпідів. Генетичні аномалії обміну сфінголіпідів-сфінголіпідози.
Сфінголіпіди - складні ліпіди біологічних мембран, що побудовані на основі високомолекулярного спирту сфінгозину. Ці ліпіди - сфінгомієліни та глікосфінголіпіди - в найбільшій кількості наявні у структурах центральної та периферичної нервової системи, зокрема в мієлінових оболонках нервів.
Для синтезу високомолекулярного аліфатичного аміноспирту сфінгозину вико-
ристовуються вуглеводневий радикал пальмітату й залишок амінокислоти ссрину.
Реакція каталізується ферментом, залежним від піридоксапьфосфату (вітаміну ВД і
потребує дії НАДФН-залежної дегідрогенази; дигідросфінгозин, що утворюється, окислюється до сфінгозину за участю специфічного флавопротеїну:
Цераміди є базовою молекулярною структурою всіх сфінголіпідів. Вони утворюються шляхом М-ацилування аміногрупи сфінгозину певною високомолекулярною жирною кислотою:
Сфінгомієліни - молекулярні структури, що утворюються шляхом приєднання
фосфохоліну до церамідів, які містять у своєму складі залишки різних жирних кислот.
Донором фосфохоліну є ЦДФ-холін.
Катаболізм сфінголіпідів
Катаболізм сфінголіпідів здійснюється шляхом послідовного розщеплення їх молекул за участю лізосомальних гідролаз.
1. Сфінгомієліни розщеплюються до цераміду та фосфохоліну за участю сфінгоміслінази:
2. Глікосфінголіпіди. Розщеплення глікосфінголіпідів починається із поступового
Найбільш поширеними є такі сфінголіпідози
Хвороба Німана - Піка: сфінголіпідоз, спричинений порушенням синтезу сфінго-
мієлінази, що супроводжується накопиченням у головному мозку, селезінці та печінці
хворих сфінгомієліну. Хвороба призводить до затримки психічного розвитку та смерті
в ранньому дитячому віці.
Хвороба Тея - Сакса - генетична хвороба, спричинена дефектом у синтезі N-ацетил-гексозамінідази, що відщеплює термінальний N-ацетилгалак тозамін від гангліозиду GМ2, який в аномальних кількостях накопичується в головному мозку. Хвороба проявляється затримкою розумового розвитку, сліпотою, неврологічними розладами, макроцефалією; смерть хворих дітей звичайно настає у віці 3-4 років.
Хвороба Гоше (глюкоцереброзидний ліпідоз) - сфінголіпідоз, генетичний дефект при якому полягає в недостатньому синтезі глюкоцереброзидази - ферменту, що відщеплює залишок глюкози від молекул глюкоцереброзидів, які накопичуються в ретикулоендотеліальній системі.
Холестерин - стероїд, що виконує важливі структурні та регуляторні функції, вхо-
дячи до складу біомембран та виступаючи попередником у синтезі фізіологічно ак-
тивних сполук різних класів. Джерелами холестерину для організму людини є його
біосинтез і надходження з продуктами харчування тваринного походження.
Усього за добу в організмі дорослої людини синтезується в середньому від 0,5 до
1,0 г холестерину, з їжею надходить 0,3-0,5 г (у деяких випадках - до 1,0-1,2 г) холе-
стерину. Здатність до синтезу холестерину мають усі клітини тваринного походження,
за винятком зрілих еритроцитів. Проте найбільша кількість ендогенного холестерину
(від 50 до 80 %) синтезується в печінці, решта стеролу утворюється в кишечнику (10-
15 %) та в шкірі (близько 5 %). Саме в клітинах печінки та слизової оболонки кишечнику стерол синтезується не тільки для власних потреб, а й “на експорт”
Попередником у біосинтезі холестерину є ацетил-КоА, який утворюється при окис-
ленні глюкози за рахунок окислювального декарбоксилування пірувату або при
β-окисленні жирних кислот.
Регуляція біосинтезу холестерину
Лімітуючим етапом у процесі біосинтезу холестерину є реакція утворення мевалонату з β-ГОМК, що каталізується β-ГОМК-редуктазою. Гальмування швидкості процесу здійсню ється за принципом негативного зворотного зв’язку, коли накопичення кінцевого продукту анаболічного шляху - холестерину - зменшує швидкість його утворення.
Інгібітором ферменту є холестерин або холестериновмісний ліпопротеїн ЛПНЩ. Відповідно до таких механізмів, споживання холестерину з їжею гальмує його
утворення в печінці, а безхолестеринова дієта, навпаки, активує ендогенний синтез холестерину в гепатоцитах.
Молекулярні механізми регуляції β-ГОМК-редуктазної реакції включають у себе
як ковалентну модифікацію ферменту (фосфорилована форма - неактивна, а де-
фосфорилована - активна), так і вплив біохімічних модуляторів на швидкість синтезу
(ферментна індукція) або деградацію ферменту.
Інсулін та гормони щитовидної залози збільшують активність β-ГОМК-редуктази, а глюкагон та глюкокортикоїди - зменшують.
64. Шляхи біотрансформації холестерину: етерифікація; утворення жовчних кислот, стероїдних гормонів, вітаміну D3.
Біотрансформація холестерину в інші біологічно активні сполуки стероїдної природи
здійснюється за рахунок введення в молекулу стеролу додаткових гідроксильних груп
та реакцій модифікації у бічному ланцюгу. Реакції окисного гідроксилу вання стероїдів
каталізуються ферментами монооксигеназами (оксигеназами мішаної функції).
Сумарне рівняння процесу окисного гідроксилування холестерину (ІІН), який відбу-
вається при синтезі його біологічно активних похідних, має вигляд:
RH + O2 + НАДФН + Н+ → R-OH +H2O+ НАДФ
Процес перебігає за участю цитохрому Р-450 у мембранах ендоплазматичного ре-
тикулуму гепатоцитів (“мікросомальне окислення”) або в мітохондріях наднирко-
вих залоз та клітин статевих залоз.
Біосинтез жовчних кислот
У гепатоцитах холестерин перетворюється на жовчні кислоти - важливі компоненти жовчі, що беруть участь у перетравлюванні харчових жирів у кишечнику людини і тварин.
Жовчні кислоти є гідроксильованими похідними холанової кислоти.
Холева та хенодезоксихолева кислоти (первинні жовчні кислоти) утворюються в
печінці при гідроксилуванні циклопентанпергідрофенантренового циклу та частко-
вому окисленні в боковому ланцюгу молекули холестерину.
Біосинтез стероїдних гормонів
Стероїдні гормони містять у своєму складі 21 (кортикоїди, прогестерон) і менше
(19 - андрогени, 18 - естрогени) атомів вуглецю, тому їх утворення з С27-стероїду
холестерину включає, крім окисного гідроксилування, реакції відновлення та ізомеризації, також розщеплення вуглеводневого бічного ланцюга.
Першим етапом на шляху синтезу з холестерину стероїдних гормонів надниркових
залоз (кортикостероїдів) є утворення С21-стероїду прегненолону - безпосереднього
попередника прогестагену прогестерону (С21), який у клітинах надниркових залоз
перетворюється на кортикостероїди (С21): глюкокортикоїд кортизол та мінералокор-
тикоїд альдостерон.
Гормони чоловічих та жіночих статевих залоз також утворюються з холестерину
через стадію прегненолону та прогестерону, який у цих органах перетворюється на
17-α-гідроксипрогестерон - попередник андрогену - тестостерону та естрогенів - естрону та естрадіолу.
Біосинтез вітаміну D3
Перетворення холестерину у вітамін D3 - холекальциферол - потребує розщеплення кільця циклопентанпергідрофенантрену з утворенням провітаміну D3, який підлягає реакціям окисного гідроксилування з утворенням біологічно активної форми вітаміну - 1,25-дигідроксихолекальциферолу (кальцитріолу).
65. Циркуляторний транспорт та депонування ліпідів у жировій тканині. Ліпопротеїнліпаза ендотелію.
Триацилгліцероли (нейтральні жири) - ліпіди, що складають основну частину
харчових ліпідів і в найбільшій кількості представлені в адипоцитах жирової тканини,
де вони виконують функцію резерву метаболічного палива. Кількість нейтральних
жирів в організмі дорослої людини масою 70 кг дорівнює в середньому 10-15 кг. Крім
жирової тканини, біосинтез триацилгліцеролів в обмеженій кількості відбувається та-
кож в інших тканинах, зокрема печінці, кишечнику, молочній залозі в період лактації.
Метаболічними попередниками в біосинтезі триацилгліцеролів є активовані жирні
кислоти (ацил-КоА) та гліцерол-3-фосфат, що, у свою чергу, постачаються за рахунок
окислення глюкози.
Особливості біосинтезу триацилгліцеролів в адипоцитах
У жировій тканині практично відсутній фермент гліцеролфосфокіназа, тому го-
ловним джерелом гліцерол-3-фосфату в адипоцитах є реакція відновлення ді-
оксіацетонфосфату, що постачається за рахунок гліколітичного розщеплення глюкози.
Таким чином, ліпогенез у жировій тканині суттєво залежить від постачання адипо-
цитів глюкозою. Оскільки проникність мембран адипоцитів для глюкози є інсулінза-
лежною, нормальне вироблення підшлунковою залозою інсуліну є важливою переду-
мовою біосинтезу триацилгліцеролів в адипоцитах.
Ліпопротеїни плазми крові
Кров людини містить декілька класів комплексів ліпідів із білками, що виконують функції міжорганного транспорту ліпідів - транспортні ліпопротеїди плазми крові. Транспортні ліпопротеїни є фізико-хімічною формою, за допомогою якої гідрофобні молекули ліпідів утримуються в стабільному стані у гідрофільному (водно-сольовому) середовищі плазми крові.
Основні класи ліпопротеїнів плазми крові'.
- хіломікрони (ХМ);
- ліпопротеїни дуже низької щільності (,ЛПДНЩ), або пре-β-ліпопротеїни;
- ліпопротеїни проміжної щільності (.ПППЩ);
- ліпопротеїни низької щільності (ЛПНЩ), або β-ліпопротеїни;
- ліпопротеїни високої щільності (ЛПВЩ), або а-ліпопротеїни.
Ці класи ліпопротеїнів розрізняються за своїми фізико-хімічними характеристиками, біохімічним (ліпідним, білковим) складом та фізіологічними функціями.
За своєю молекулярною будовою ліпопротеїни плазми крові - це кулясті структури -
міцели, всередині яких міститься гідрофобна ліпідна серцевина (ядро), що складається переважно з триацилгліцеролів та ефірів холестерину. Гідрофобне ядро вкрите
шаром полярних амфіпатичних фосфоліпідів, периферичних та інтегральних білків.
Гіперліпопротеїнемії
Гіперліпопротеїнемія - клініко-біохімічний синдром, при якому в плазмі крові людини спостерігається підвищення (порівняно з нормою для певної популяції) концентрації певних класів ліпопротеїнів, а також триацилгліцеролів та холестерину.
За механізмом походження виділяють:
- первинні (спадкові) гіперліпопротеїнемії, тобто такі, що спричинені генетичними дефектами в синтезі певних ферментів обміну ліпідів крові (зокрема, ліпопротеїн-
ліпази, холестсрол-ацилтрансферази) або неферментних білків - порушеннями в син-
тезі певних апопротеїнів, рецепторів дія апобілків та ліпопротеїнів (зокрема ЛПНЩ);
- вторинні (набуті) гіперліпопротеїнемії- гіперліпопротеїнемії, що розвиваються
внаслідок певних хвороб внутрішніх органів (гепатит, цироз печінки, нефроз), ендо-
кринопатій (порушення функції щитовидної залози, статевих залоз, цукровий діабет),
дії пошкоджуючих факторів середовища (хронічний алкоголізм).
Атеросклероз
Атеросклероз - хвороба, головним проявом якої є відкладання в судинних стінках
ліпідних утворень - “бляшок”, основними біохімічними компонентами яких є холестерин та його ефіри. Навкруги ліпідних бляшок в інтимі судин виникає клітинна реакція, що включає в себе утворення фіброзної тканини та проліферацію їладком’язових клітин. Атеросклеротичні бляшки спричиняють звуження кровоносних судин, посилене згортання крові в ділянках їх локалізації та, як результат, порушення кровопостачання відповідних органів і тканин. Як наслідок атеросклерозу розвиваються ішемічна хвороба серця, інфаркт міокарда й порушення церебрального кровообігу, що стають важливою причиною смерті людей дорослого та похилого віку.
Біохімічною основою розвитку атеросклерозу є підвищена концентрацій в крові людини холестерину - гіперхолестеринемія, спричинена різними факторами - дієтарними, ендокринними, генетичними.
Ожиріння
Ожиріння - стан, що характеризується надмірним накопиченням у жировій тканині триацилгліцеролів. При ожирінні збільшується кількість жирових клітин (адипоцитів) або їх розмір. Загальна маса нейтральних жирів в організмі людини за умов
ожиріння може досягати значних кількостей. Порушення ліпідного обміну, що відбувається при ожирінні, часто поєднуються з наявністю у хворого атеросклерозу або цукрового діабету.
Цукровий діабет
Цукровий діабет традиційно розглядається як патологія, що первинно пов’язана
з порушеннями вуглеводного обміну. Найбільш характерним біохімічним проявом різних типів цукрового діабету в клініці є гіперглікемія (гіперглюкоземія), яка
розвивається внаслідок втрати специфічного впливу інсуліну на проникність клітинних мембран дія глюкози. Але метаболічні ефекти інсуліну поширюються на багато аспектів обміну глюкози, ліпідів та амінокислот, у зв’язку з чим цукровий діабет є хворобою, при якому відбуваються глибокі порушення не тільки вуглеводного, але й ліпідного та білкового обмінів.
Розрізняють інсулінозалежний цукровий діабет (діабет І типу, ювенільний діабет) та інсулінонезалежний цукровий діабет (діабет II типу, діабет похилого віку).
68. Пул вільних амінокислот в організмі: шляхи надходження та використання вільних амінокислот в тканинах.
Кількість амінокислот, що перетворюються за добу, складає в організмі дорослої здорової людини в стані азотистої рівноваги 300-500 г, а стаціонарна їх концентрація дорівнює близько 50-100 г на масу тіла.
Потік амінокислот що входить в організм з амінокислотного пулу складається з таких джерел:
1. Амінокислот, які всмоктуються ентероцитами кишечника внаслідок гідролізу
харчових білків у травному каналі
2. Амінокислот, які вивільняються в результаті розщеплення власних клітинних і
позаклітинних білків.
3. Амінокислот, які синтезуються в організмі.
Потік амінокислот, що виходить з амінокислотного пулу, включає анаболічні і
катаболічні шляхи перетворення вільних амінокислот і складається з таких компонентів:
1. Використання амінокислот для синтезу білків організму.
2. Використання амінокислот, які не включені в анаболічні процеси, в катаболічних
реакціях.
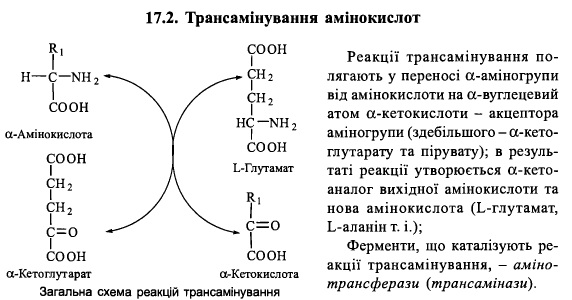
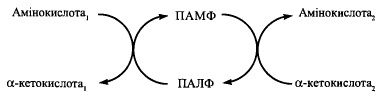
69. Трансамінування амінокислот: реакції та їх біохімічне значення, механізми дії амінотрансфераз.
Амінотрансферазні реакції
У різних тканинах організму людини і тварин міститься більше десяти різних амінотрансфераз, що розрізняються за своєю субстратною специфічністю.
Найбільш поширеними є такі амінотрансферази:
(1) аланінамінотрансфераза (глутамат-піруваттрансаміназа - ГПТ):
L-аланін + а-кетоглутарат = Піруват + L-глутамат
(2) аспартатамінотрансфераза (глутамат-оксалоацетаттрансаміназа - ГОТ):
L-аспартат + а-кетоглутарат = Оксалоацетат + L-глутамат
(3) тирозинамінотрансфераза:
L-тирозин + а-кетоглутарат = р-гідроксифенілпіруват + L-глутамат
(4) лейцинамінотрансфераза:
L-лейцин + а-кетоглутарат = а-кетоізокапроат + L-глутамат
Реакції трансамінування, що каталізуються амінотрансферазами, активно перебігають в багатьох органах, найактивніше - в печінці, скелетних м’язах, міокарді, головному мозку, нирках. Визначення активності аланіналйнотрансферази (аламінової трансамінази - АлАТ) та аспартатамінотрансферази (аспарагінової трансамінази - АсАТ) широко застосовується в медичній практиці з мстою діагностики пошкоджень внутрішніх органів. Внаслідок виходу цих ферментних білків через ушкоджені клітинні мембрани в кров при інфаркті міокарда спостерігається значне
підвищення активності в сироватці крові АсАТ, при вірусних та токсичних пошкодженнях печінки - АлАТ.
Механізм дії
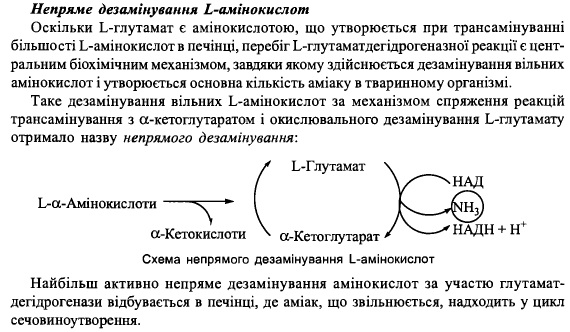
70. Пряме та непряме дезамінування вільних L-амінокислот в тканинах.
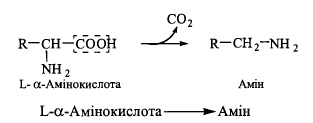
71. Декарбоксилювання L -амінокислот в організмі людини. Фізіологічне значення утворених продуктів. Окислення біогенних амінів.
Реакція дскарбоксилування амінокислот полягає у відщепленні діоксиду вуглецю
від молекули амінокислоти з утворенням амінів (біогенних амінів), значна частина
яких має високу фізіологічну активність як гормони, нейромедіатори, або є їх попередниками чи метаболітами:
Реакція каталізується ферментами - декарбоксилазами амінокислот, коферментом яких є піридоксальфосфат, що в ході каталітичного акту утворює з амінокислотами шифові основи, подібні до розглянутих в реакціях транс амінування. Декарбоксилази амінокислот є стереоспецифічними ферментами, що діють тільки на L-стереоізомери.
Дскарбоксилуванню можуть підлягати як ациклічні, так і циклічні амінокислоти; в сечі людини знайдено близько сорока різних біогенних амінів.
Процеси декарбоксилування амінокислот активно перебігають у порожнині товстої кишки під дією ферментів мікроорганізмів, що є компонентами нормальної мікрофлори травного тракту людини (“бактеріальне гниття білків у кишечнику”).
Прикладами реакцій декарбоксилування амінокислот у кишечнику є утворення
токсичних діамінів, т.з. птомаїнів (“трупних отрут”) з діаміномонокарбонових кислот орнітину та путресцину.
Зокрема, продуктом декарбоксилування орнітину (а, 8-діаміновалсріанової кислоти) є тетраметилендіамін путресцин, а продуктом декарбоксилування лізину (а, є діамінокапронової кислоти) - пентаметилендіамін кадаверин.
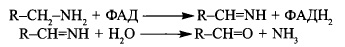
Окислення біогенних амінів
Накопичення біогенних амінів в організмі спричиняє несприятливі патофізіологічні
зміни з боку серцево-судинної системи, кишечника, інших гладком’язових органів.
Знешкодження (детоксикація) фізіологічно активних амінів відбувається в клітинах
печінки за участі моноамінооксидази мітохондрій - ФАД-залежного ферменту, що
спричиняє окислювальне дезамінування амінів до альдегідів:
Альдегіди - продукти дезамінування біогенних амінів - окислюються до відповідних
кислот і підлягають подальшій окислювальній деградації або екскретуються з організму із сечею. Аміак надходить у систему синтезу сечовини.
72. Шляхи утворення та знешкодження аміаку в організмі.
Головним у кількісному відношенні джерелом наконичення аміаку в організмі людини є окислювальне дезамінування амінокислот, тобто білковий катаболізм: азот сечовини - кінцевого азотовмісного продукту дсградацї білків – складає близько 90 % всього азоту, що екскретується. Додатковими джерелами ендогенного аміаку є реакції дезамінування біогенних амінів, азотистих основ, які утворюються при катаболізмі нуклеотидів. Значна кількість вільного аміаку всмоктується в кров із системи
ворітної вени (v. porta) внаслідок його утворення при катаболізмі азотовмісних біоорганічних сполук (головним чином, білків продуктів харчування) кишковими бактеріями.
Основним джерелом утворення аміаку в тканині головного мозку є реакція гідролітичного дезамінування АМФ до інозинмонофосфату (ІМФ), що каталізується ферментом аденозиндезаміназою:
АМФ+H2O --- ІМФ+NH3
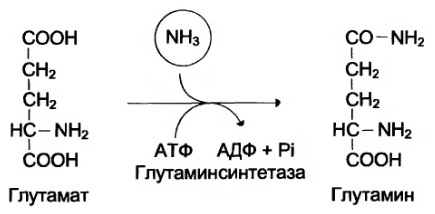
Аміак, що вивільняється, знешкоджується в результаті глуглутамінсинтетазної реакції, утворюючи з L-глутамату глутамін, який виводиться з головного мозку.
Основной реакцией связывания аммиака, протекающей во всех тканях организма, явля ется синтез глутамина под действием глутаминсинтетазы:
Ещё одной реакцией обезвреживания аммиака в тканях можно считать синтез аспарагина под действием аспарагинсинтетазы.
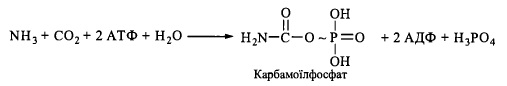
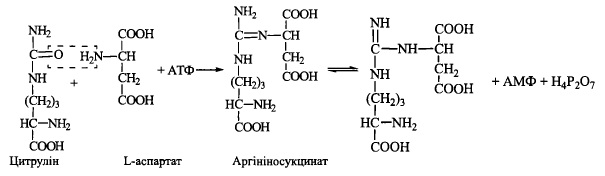
1. Утворення з аміаку та дюксиду вуглецю за участю АТФ карбамоїлфосфату:
Реакція каталізується карбамоїлфосфатсинтетазою. Джерелом аміногрупи (у вигляді молекули аміаку) с глутаматдегідрогсназна реакція.
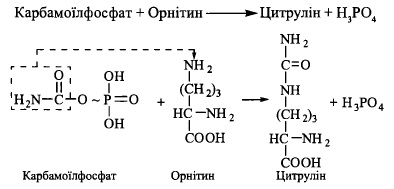
2. Перенесення карбамоїльної групи на орнітин з утворенням цитруліну (фермент -
орнітинкарбамоїлтрансфераза):
3. Акцептування другої аміногрупи шляхом взаємодії цитруліну з L-аспартатом
(фермент - аргініносукцинатсинтетаза):
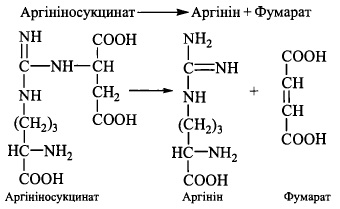
4. Розщеплення аргініносукцинату при дії ферменту аргініносукцинатліази; продуктами реакції є: аргінін - безпосередній попередник сечовини - та фумарат:
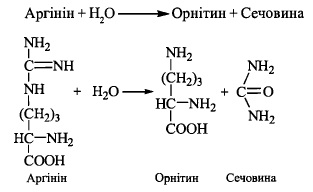
5. Гідроліз аргініну при дії ферменту аргінази з утворенням сечовини та регенера-
цією орнітину (завершення метаболічного циклу):
Генетичні дефекти ферментів синтезу сечовини
Існують спадкові ензимопатії, спричинені повним або частковим дефектом утворення в печінці окремих ферментів циклу сечовиноутворення. Найважчими клінічними проявами характеризуються порушення синтезу карбамоїлфосфатсинтетази
та орнітинкарбамоїлтрансферази. Діти з такими генетичними дефектами страждають вираженою енцефалопатією, прояви якої дещо послаблюються в умовах повного виключення споживання харчових білків.
74. Загальні шляхи метаболізму вуглецевих скелетів амінокислот в організмі людини. Глюкогенні та кетогенні амінокислоти.
Двадцять L-амінокислот, що розрізняються за своєю хімічною структурою, біоло-
гічною роллю та особливостями метаболізму, входять до складу білків організму і
присутні в клітинах та екстрацелюлярних просторах у вільному стані. Безазотисті
скелети вільних амінокислот, які утворюються в результаті трансаміпувашія та де-
замінування, - це метаболіти гліколізу, цитратного циклу, Р-окислення жирних кислот
або речовини, що можуть перетворюватися в інтермедіати цих головних катаболіч-
них шляхів організму.
Глюкогенні амінокислоти
L-Амінокислоти, що метаболізуються в циклі трикарбонових кислот, можуть включати свої вуглецеві скелети в молекули глюкози. Ці амінокислоти, використання яких у синтезі глюкози реалізується після їх входження в ЦТК через ацетил-КоА,
а-кетоглутарат, сукциніл-КоА та фумарат, отримали назву глюкогенних амінокислот.
Кетогенні амінокислоти
Дві L-амінокислоти включаються в катаболізм тільки через ацетоацстил-КоА, який
у клітинах печінки може перетворюватися на кетонові тіла ацетоацетат та
β-гідроксибутират. Це - кетогенні амінокислоти. Деякі амінокислоти віддають свої
вуглецеві фрагменти на утворення як глюкози, так і кетонових тіл.
Кетогенез із амінокислот має особливо негативне значення при деяких порушеннях
ферментних процесів, зокрема при некомпенсованому цукровому діабеті, у зв’язку з
чим таким хворим рекомендується обмежувати надходження кетогенних амінокис-
лот у складі продуктів харчування.
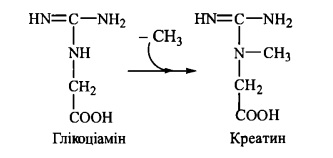
75. Біосинтез та біологічна роль креатину і креатинфосфату.
Креатин - азотиста сполука, яка у вигляді креатинфосфату має важливе значення в енергозабезпеченні функції м’язів.
Біосинтез креатину відбувається за участю амінокислот гліцину, аргініну та метіо-
ніну. Процес синтезу складається з двох стадій:
1-ша стадія - відбувається в нирках і полягає в утворенні глікоціаміну (гуанідинацетату) із аргініну та гліцину (фермент гліцинамідинотрансфераза)'.
2-а стадія - відбувається в печінці, куди тікоціамін надходить з током крові, і полягає в метилюванні глікоціаміну до креатину за участю 8-аденозилметіоніну (фермент гуанідинацетатметилтрансфераза):
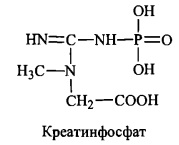
Фосфорилування креатину при дії креатинфосфокінази генерує креатинфосфат - джерело термінової регенерації АТФ при м’язовому скороченні. Незворотна
нсфермснтативна дегідратація і дефосфорилування крсатинфосфату призводить до
утворення ангідриду креатину - креатиніну.
У формі креатиніну з організму людини виділяється із сечею значна частина азоту
амінокислот; у здорової людини виділення креатиніну пропорційне масі м’язових тканин і значно збільшується за умов травматичних пошкоджень м’язів.
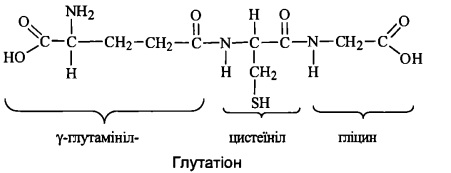
76. Глутатіон: будова, біосинтез та біологічні функції глутатіону
Глутатіон - трипептид γ-глутамініл-цистеїннл-гліцин, що має у своєму складі вільну
сульфгідрильну групу:
Глутатіон міститься в клітинах тваринного організму у високій концентрації (близько
5 мМ). Глутатіон оборотно перетворюється з відновленої (Г-SН) до окисленої
(Г-S-S-Г) форми, відіграючи роль буфера SН-груп.
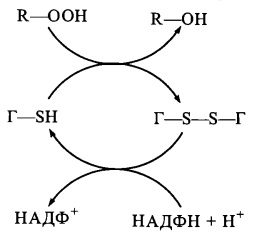
Біохімічна функція глутатіону в організмі пов’язана з відновленням і детоксикацією органічних пероксидів - похідних пероксиду водню НО-ОН, у молекулі якого один (гідропероксиди) або обидва (алкілпероксиди) атоми водню заміщені на алкільні радикали:
R-O-O-H R-O-O-R
При взаємодії глутатіону з гідропероксидом утворюються нешкідливі органічні спир-
ти, що підлягають подальшому окисленню:
Реакція каталізується ферментом глутатіонпероксидазою, що містить в актив-
ному центрі атом селену (Sе).
Зворотне відновлення Г-SS-Г до Г-SH каталізується НАДФН-залежною глутатіонредуктазою.
77. Спеціалізовані шляхи метаболізму циклічних амінокислот - фенілаланіну, та тирозину.
Особливістю метаболізму в тваринних організмах циклічних амінокислот феніла-
ланіну та тирозину є утворення з них численних фізіологічно активних сполук гормо-
нальної та медіаторної дії, а саме: катехоламінів (адреналіну, норадреналіну), тиреоїд-
них гормонів, меланінів (рис. 18.7).
1. Шляхи метаболізму фенілаланіну
1.1. Катаболічний шлях обміну полягає у втраті фенілаланіном аміногрупи (в ре-
акції трансамінування) з утворенням фенілпірувату та кінцевого метаболіту фенілаце-
тату, що екскретується з організму.
1.2. Шлях синтезу фізіологічно активних сполук починається з перетворення фе-
нілаланіну на тирозин при дії ферменту фенілаланіпгідроксилази з подальшим пере-
творенням тирозину (див. нижче).
2. Шляхи метаболізму тирозину
2.1. Катаболічний шлях обміну полягає у трансамінуванні тирозину і перетворенні
його на р-оксифенілпіруват, який окислюється до гомогентизинової кислоти у склад-
ній реакції, коферменту роль у якій виконує аскорбінова кислота (вітамін С); подальші
перетворення полягають в окисленні гомогентизату до фумарилацетоацетату (фермент оксидаза гомогентизинової кислоти) та розщепленні фумарилацетоацетату
до фумарату та ацетоацетату.
2.2. Шлях синтезу катехоламінів та меланінів (пігментів шкіри). Шлях починається
з окислення тирозину за участю специфічної гідроксилази до 3,4-діоксифенілаланіну
(ДОФА), на рівні якого відбувається дивергенція двох обмінних шляхів: утворення
катехоламінів (через декарбоксилування до дофаміну) та меланінів (через окислення
тирозиназою до дофахінону).
2.3. Шлях синтезу тиреоїдних гормонів-реалізується в клітинах щитовидної зало-
зи і полягає в утворенні йодованих тиронінів.
78. Спадкові ензимопатії обміну циклічних амінокислот - фенілаланіну та тирозину.
studopedia.su - Студопедия (2013 - 2025) год. Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав!Последнее добавление
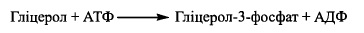 1.2. Шляхом відновлення діоксіацетонфосфату - інтермедіату гліколітнчного роз-
іцсшісшія глюкози. Реакція каталізується НАДН-залежною гліцерол-3-фосфат-де-
гідрогеназою (а-гліцерофосфатдегідрогеназою):
1.2. Шляхом відновлення діоксіацетонфосфату - інтермедіату гліколітнчного роз-
іцсшісшія глюкози. Реакція каталізується НАДН-залежною гліцерол-3-фосфат-де-
гідрогеназою (а-гліцерофосфатдегідрогеназою):
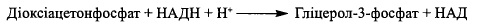 Гліцерофосфоліпіди (фосфогліцериди) - фосфатидилхолін, фосфатидил-етаноламін, фосфати дилсерин і кардіол іній - належать до структурних ліпідів, що складають ліпідний матрикс біологічних мембран. Це - складні ліпіди, побудовані на основі гліцеролу, тому перші етапи їх біосинтезу однакові з розглянутими вище фермен-
тативними реакціями утворення триацилгліцеролів, а саме:
Гліцерол → Гліцерол-З-фосфат → Фосфатидна кислота
Після утворення 1,2 діацилптіцеролу реакції синтезу триацилгліцеролів та фосфогліцеридів дивергують.
У разі біосинтезу зазначених фосфогліцеридів до 1,2-діацилгліцеролу, що утворюєть-
ся в результаті гідролізу фосфатидної кислоти, приєднується гідрофільна головка, яка
містить аміноспирт (холін або етаноламін). Особливістю процесу є використання в
реакції активованих форм аміноспиртів - комплексів холіну (етаноламіну) з нуклео-
зиддифосфатом ЦДФ, які утворюються за рахунок таких реакцій:
- активації холіну (етаноламіну) шляхом АТФ-залежного фосфорилування аміно-
спирту;
- взаємодії фосфохоліну (або фосфоетаноламіну) з нуклеозидтрифосфатом ЦТФ з
утворенням ЦДФ-холіну (ЦДФ-етаноламіну);
- При взаємодії ЦДФ-холіну (ЦДФ-етаноламіну) з 1,2-діацилгліцеролом утворю-
ються фосфатидилхолін або фосфатидилетаноламін.
Гліцерофосфоліпіди (фосфогліцериди) - фосфатидилхолін, фосфатидил-етаноламін, фосфати дилсерин і кардіол іній - належать до структурних ліпідів, що складають ліпідний матрикс біологічних мембран. Це - складні ліпіди, побудовані на основі гліцеролу, тому перші етапи їх біосинтезу однакові з розглянутими вище фермен-
тативними реакціями утворення триацилгліцеролів, а саме:
Гліцерол → Гліцерол-З-фосфат → Фосфатидна кислота
Після утворення 1,2 діацилптіцеролу реакції синтезу триацилгліцеролів та фосфогліцеридів дивергують.
У разі біосинтезу зазначених фосфогліцеридів до 1,2-діацилгліцеролу, що утворюєть-
ся в результаті гідролізу фосфатидної кислоти, приєднується гідрофільна головка, яка
містить аміноспирт (холін або етаноламін). Особливістю процесу є використання в
реакції активованих форм аміноспиртів - комплексів холіну (етаноламіну) з нуклео-
зиддифосфатом ЦДФ, які утворюються за рахунок таких реакцій:
- активації холіну (етаноламіну) шляхом АТФ-залежного фосфорилування аміно-
спирту;
- взаємодії фосфохоліну (або фосфоетаноламіну) з нуклеозидтрифосфатом ЦТФ з
утворенням ЦДФ-холіну (ЦДФ-етаноламіну);
- При взаємодії ЦДФ-холіну (ЦДФ-етаноламіну) з 1,2-діацилгліцеролом утворю-
ються фосфатидилхолін або фосфатидилетаноламін.
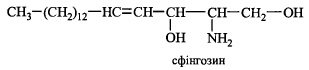 Для синтезу високомолекулярного аліфатичного аміноспирту сфінгозину вико-
ристовуються вуглеводневий радикал пальмітату й залишок амінокислоти ссрину.
Реакція каталізується ферментом, залежним від піридоксапьфосфату (вітаміну ВД і
потребує дії НАДФН-залежної дегідрогенази; дигідросфінгозин, що утворюється, окислюється до сфінгозину за участю специфічного флавопротеїну:
Цераміди є базовою молекулярною структурою всіх сфінголіпідів. Вони утворюються шляхом М-ацилування аміногрупи сфінгозину певною високомолекулярною жирною кислотою:
Сфінгомієліни - молекулярні структури, що утворюються шляхом приєднання
фосфохоліну до церамідів, які містять у своєму складі залишки різних жирних кислот.
Донором фосфохоліну є ЦДФ-холін.
Катаболізм сфінголіпідів
Катаболізм сфінголіпідів здійснюється шляхом послідовного розщеплення їх молекул за участю лізосомальних гідролаз.
1. Сфінгомієліни розщеплюються до цераміду та фосфохоліну за участю сфінгоміслінази:
2. Глікосфінголіпіди. Розщеплення глікосфінголіпідів починається із поступового
Найбільш поширеними є такі сфінголіпідози
Хвороба Німана - Піка: сфінголіпідоз, спричинений порушенням синтезу сфінго-
мієлінази, що супроводжується накопиченням у головному мозку, селезінці та печінці
хворих сфінгомієліну. Хвороба призводить до затримки психічного розвитку та смерті
в ранньому дитячому віці.
Хвороба Тея - Сакса - генетична хвороба, спричинена дефектом у синтезі N-ацетил-гексозамінідази, що відщеплює термінальний N-ацетилгалак тозамін від гангліозиду GМ2, який в аномальних кількостях накопичується в головному мозку. Хвороба проявляється затримкою розумового розвитку, сліпотою, неврологічними розладами, макроцефалією; смерть хворих дітей звичайно настає у віці 3-4 років.
Хвороба Гоше (глюкоцереброзидний ліпідоз) - сфінголіпідоз, генетичний дефект при якому полягає в недостатньому синтезі глюкоцереброзидази - ферменту, що відщеплює залишок глюкози від молекул глюкоцереброзидів, які накопичуються в ретикулоендотеліальній системі.
Для синтезу високомолекулярного аліфатичного аміноспирту сфінгозину вико-
ристовуються вуглеводневий радикал пальмітату й залишок амінокислоти ссрину.
Реакція каталізується ферментом, залежним від піридоксапьфосфату (вітаміну ВД і
потребує дії НАДФН-залежної дегідрогенази; дигідросфінгозин, що утворюється, окислюється до сфінгозину за участю специфічного флавопротеїну:
Цераміди є базовою молекулярною структурою всіх сфінголіпідів. Вони утворюються шляхом М-ацилування аміногрупи сфінгозину певною високомолекулярною жирною кислотою:
Сфінгомієліни - молекулярні структури, що утворюються шляхом приєднання
фосфохоліну до церамідів, які містять у своєму складі залишки різних жирних кислот.
Донором фосфохоліну є ЦДФ-холін.
Катаболізм сфінголіпідів
Катаболізм сфінголіпідів здійснюється шляхом послідовного розщеплення їх молекул за участю лізосомальних гідролаз.
1. Сфінгомієліни розщеплюються до цераміду та фосфохоліну за участю сфінгоміслінази:
2. Глікосфінголіпіди. Розщеплення глікосфінголіпідів починається із поступового
Найбільш поширеними є такі сфінголіпідози
Хвороба Німана - Піка: сфінголіпідоз, спричинений порушенням синтезу сфінго-
мієлінази, що супроводжується накопиченням у головному мозку, селезінці та печінці
хворих сфінгомієліну. Хвороба призводить до затримки психічного розвитку та смерті
в ранньому дитячому віці.
Хвороба Тея - Сакса - генетична хвороба, спричинена дефектом у синтезі N-ацетил-гексозамінідази, що відщеплює термінальний N-ацетилгалак тозамін від гангліозиду GМ2, який в аномальних кількостях накопичується в головному мозку. Хвороба проявляється затримкою розумового розвитку, сліпотою, неврологічними розладами, макроцефалією; смерть хворих дітей звичайно настає у віці 3-4 років.
Хвороба Гоше (глюкоцереброзидний ліпідоз) - сфінголіпідоз, генетичний дефект при якому полягає в недостатньому синтезі глюкоцереброзидази - ферменту, що відщеплює залишок глюкози від молекул глюкоцереброзидів, які накопичуються в ретикулоендотеліальній системі.
 Інгібітором ферменту є холестерин або холестериновмісний ліпопротеїн ЛПНЩ. Відповідно до таких механізмів, споживання холестерину з їжею гальмує його
утворення в печінці, а безхолестеринова дієта, навпаки, активує ендогенний синтез холестерину в гепатоцитах.
Молекулярні механізми регуляції β-ГОМК-редуктазної реакції включають у себе
як ковалентну модифікацію ферменту (фосфорилована форма - неактивна, а де-
фосфорилована - активна), так і вплив біохімічних модуляторів на швидкість синтезу
(ферментна індукція) або деградацію ферменту.
Інсулін та гормони щитовидної залози збільшують активність β-ГОМК-редуктази, а глюкагон та глюкокортикоїди - зменшують.
Інгібітором ферменту є холестерин або холестериновмісний ліпопротеїн ЛПНЩ. Відповідно до таких механізмів, споживання холестерину з їжею гальмує його
утворення в печінці, а безхолестеринова дієта, навпаки, активує ендогенний синтез холестерину в гепатоцитах.
Молекулярні механізми регуляції β-ГОМК-редуктазної реакції включають у себе
як ковалентну модифікацію ферменту (фосфорилована форма - неактивна, а де-
фосфорилована - активна), так і вплив біохімічних модуляторів на швидкість синтезу
(ферментна індукція) або деградацію ферменту.
Інсулін та гормони щитовидної залози збільшують активність β-ГОМК-редуктази, а глюкагон та глюкокортикоїди - зменшують.
 Амінотрансферазні реакції
У різних тканинах організму людини і тварин міститься більше десяти різних амінотрансфераз, що розрізняються за своєю субстратною специфічністю.
Найбільш поширеними є такі амінотрансферази:
(1) аланінамінотрансфераза (глутамат-піруваттрансаміназа - ГПТ):
L-аланін + а-кетоглутарат = Піруват + L-глутамат
(2) аспартатамінотрансфераза (глутамат-оксалоацетаттрансаміназа - ГОТ):
L-аспартат + а-кетоглутарат = Оксалоацетат + L-глутамат
(3) тирозинамінотрансфераза:
L-тирозин + а-кетоглутарат = р-гідроксифенілпіруват + L-глутамат
(4) лейцинамінотрансфераза:
L-лейцин + а-кетоглутарат = а-кетоізокапроат + L-глутамат
Реакції трансамінування, що каталізуються амінотрансферазами, активно перебігають в багатьох органах, найактивніше - в печінці, скелетних м’язах, міокарді, головному мозку, нирках. Визначення активності аланіналйнотрансферази (аламінової трансамінази - АлАТ) та аспартатамінотрансферази (аспарагінової трансамінази - АсАТ) широко застосовується в медичній практиці з мстою діагностики пошкоджень внутрішніх органів. Внаслідок виходу цих ферментних білків через ушкоджені клітинні мембрани в кров при інфаркті міокарда спостерігається значне
підвищення активності в сироватці крові АсАТ, при вірусних та токсичних пошкодженнях печінки - АлАТ.
Механізм дії
Амінотрансферазні реакції
У різних тканинах організму людини і тварин міститься більше десяти різних амінотрансфераз, що розрізняються за своєю субстратною специфічністю.
Найбільш поширеними є такі амінотрансферази:
(1) аланінамінотрансфераза (глутамат-піруваттрансаміназа - ГПТ):
L-аланін + а-кетоглутарат = Піруват + L-глутамат
(2) аспартатамінотрансфераза (глутамат-оксалоацетаттрансаміназа - ГОТ):
L-аспартат + а-кетоглутарат = Оксалоацетат + L-глутамат
(3) тирозинамінотрансфераза:
L-тирозин + а-кетоглутарат = р-гідроксифенілпіруват + L-глутамат
(4) лейцинамінотрансфераза:
L-лейцин + а-кетоглутарат = а-кетоізокапроат + L-глутамат
Реакції трансамінування, що каталізуються амінотрансферазами, активно перебігають в багатьох органах, найактивніше - в печінці, скелетних м’язах, міокарді, головному мозку, нирках. Визначення активності аланіналйнотрансферази (аламінової трансамінази - АлАТ) та аспартатамінотрансферази (аспарагінової трансамінази - АсАТ) широко застосовується в медичній практиці з мстою діагностики пошкоджень внутрішніх органів. Внаслідок виходу цих ферментних білків через ушкоджені клітинні мембрани в кров при інфаркті міокарда спостерігається значне
підвищення активності в сироватці крові АсАТ, при вірусних та токсичних пошкодженнях печінки - АлАТ.
Механізм дії
 Реакція каталізується ферментами - декарбоксилазами амінокислот, коферментом яких є піридоксальфосфат, що в ході каталітичного акту утворює з амінокислотами шифові основи, подібні до розглянутих в реакціях транс амінування. Декарбоксилази амінокислот є стереоспецифічними ферментами, що діють тільки на L-стереоізомери.
Дскарбоксилуванню можуть підлягати як ациклічні, так і циклічні амінокислоти; в сечі людини знайдено близько сорока різних біогенних амінів.
Процеси декарбоксилування амінокислот активно перебігають у порожнині товстої кишки під дією ферментів мікроорганізмів, що є компонентами нормальної мікрофлори травного тракту людини (“бактеріальне гниття білків у кишечнику”).
Прикладами реакцій декарбоксилування амінокислот у кишечнику є утворення
токсичних діамінів, т.з. птомаїнів (“трупних отрут”) з діаміномонокарбонових кислот орнітину та путресцину.
Зокрема, продуктом декарбоксилування орнітину (а, 8-діаміновалсріанової кислоти) є тетраметилендіамін путресцин, а продуктом декарбоксилування лізину (а, є діамінокапронової кислоти) - пентаметилендіамін кадаверин.
Окислення біогенних амінів
Накопичення біогенних амінів в організмі спричиняє несприятливі патофізіологічні
зміни з боку серцево-судинної системи, кишечника, інших гладком’язових органів.
Знешкодження (детоксикація) фізіологічно активних амінів відбувається в клітинах
печінки за участі моноамінооксидази мітохондрій - ФАД-залежного ферменту, що
спричиняє окислювальне дезамінування амінів до альдегідів:
Реакція каталізується ферментами - декарбоксилазами амінокислот, коферментом яких є піридоксальфосфат, що в ході каталітичного акту утворює з амінокислотами шифові основи, подібні до розглянутих в реакціях транс амінування. Декарбоксилази амінокислот є стереоспецифічними ферментами, що діють тільки на L-стереоізомери.
Дскарбоксилуванню можуть підлягати як ациклічні, так і циклічні амінокислоти; в сечі людини знайдено близько сорока різних біогенних амінів.
Процеси декарбоксилування амінокислот активно перебігають у порожнині товстої кишки під дією ферментів мікроорганізмів, що є компонентами нормальної мікрофлори травного тракту людини (“бактеріальне гниття білків у кишечнику”).
Прикладами реакцій декарбоксилування амінокислот у кишечнику є утворення
токсичних діамінів, т.з. птомаїнів (“трупних отрут”) з діаміномонокарбонових кислот орнітину та путресцину.
Зокрема, продуктом декарбоксилування орнітину (а, 8-діаміновалсріанової кислоти) є тетраметилендіамін путресцин, а продуктом декарбоксилування лізину (а, є діамінокапронової кислоти) - пентаметилендіамін кадаверин.
Окислення біогенних амінів
Накопичення біогенних амінів в організмі спричиняє несприятливі патофізіологічні
зміни з боку серцево-судинної системи, кишечника, інших гладком’язових органів.
Знешкодження (детоксикація) фізіологічно активних амінів відбувається в клітинах
печінки за участі моноамінооксидази мітохондрій - ФАД-залежного ферменту, що
спричиняє окислювальне дезамінування амінів до альдегідів:
 Альдегіди - продукти дезамінування біогенних амінів - окислюються до відповідних
кислот і підлягають подальшій окислювальній деградації або екскретуються з організму із сечею. Аміак надходить у систему синтезу сечовини.
Альдегіди - продукти дезамінування біогенних амінів - окислюються до відповідних
кислот і підлягають подальшій окислювальній деградації або екскретуються з організму із сечею. Аміак надходить у систему синтезу сечовини.
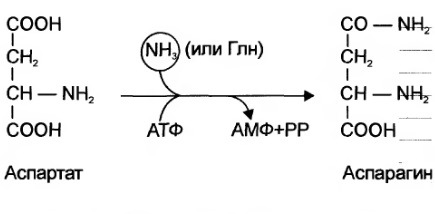
 Ещё одной реакцией обезвреживания аммиака в тканях можно считать синтез аспарагина под действием аспарагинсинтетазы.
Ещё одной реакцией обезвреживания аммиака в тканях можно считать синтез аспарагина под действием аспарагинсинтетазы.
 2. Перенесення карбамоїльної групи на орнітин з утворенням цитруліну (фермент -
орнітинкарбамоїлтрансфераза):
2. Перенесення карбамоїльної групи на орнітин з утворенням цитруліну (фермент -
орнітинкарбамоїлтрансфераза):
 3. Акцептування другої аміногрупи шляхом взаємодії цитруліну з L-аспартатом
(фермент - аргініносукцинатсинтетаза):
3. Акцептування другої аміногрупи шляхом взаємодії цитруліну з L-аспартатом
(фермент - аргініносукцинатсинтетаза):
 4. Розщеплення аргініносукцинату при дії ферменту аргініносукцинатліази; продуктами реакції є: аргінін - безпосередній попередник сечовини - та фумарат:
4. Розщеплення аргініносукцинату при дії ферменту аргініносукцинатліази; продуктами реакції є: аргінін - безпосередній попередник сечовини - та фумарат:
 5. Гідроліз аргініну при дії ферменту аргінази з утворенням сечовини та регенера-
цією орнітину (завершення метаболічного циклу):
5. Гідроліз аргініну при дії ферменту аргінази з утворенням сечовини та регенера-
цією орнітину (завершення метаболічного циклу):
 Генетичні дефекти ферментів синтезу сечовини
Існують спадкові ензимопатії, спричинені повним або частковим дефектом утворення в печінці окремих ферментів циклу сечовиноутворення. Найважчими клінічними проявами характеризуються порушення синтезу карбамоїлфосфатсинтетази
та орнітинкарбамоїлтрансферази. Діти з такими генетичними дефектами страждають вираженою енцефалопатією, прояви якої дещо послаблюються в умовах повного виключення споживання харчових білків.
Генетичні дефекти ферментів синтезу сечовини
Існують спадкові ензимопатії, спричинені повним або частковим дефектом утворення в печінці окремих ферментів циклу сечовиноутворення. Найважчими клінічними проявами характеризуються порушення синтезу карбамоїлфосфатсинтетази
та орнітинкарбамоїлтрансферази. Діти з такими генетичними дефектами страждають вираженою енцефалопатією, прояви якої дещо послаблюються в умовах повного виключення споживання харчових білків.
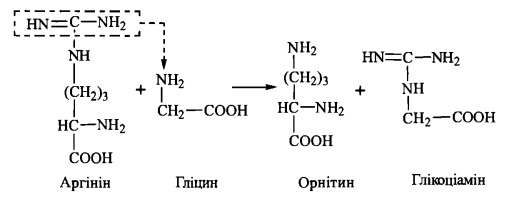 2-а стадія - відбувається в печінці, куди тікоціамін надходить з током крові, і полягає в метилюванні глікоціаміну до креатину за участю 8-аденозилметіоніну (фермент гуанідинацетатметилтрансфераза):
2-а стадія - відбувається в печінці, куди тікоціамін надходить з током крові, і полягає в метилюванні глікоціаміну до креатину за участю 8-аденозилметіоніну (фермент гуанідинацетатметилтрансфераза):
 Фосфорилування креатину при дії креатинфосфокінази генерує креатинфосфат - джерело термінової регенерації АТФ при м’язовому скороченні. Незворотна
Фосфорилування креатину при дії креатинфосфокінази генерує креатинфосфат - джерело термінової регенерації АТФ при м’язовому скороченні. Незворотна
 нсфермснтативна дегідратація і дефосфорилування крсатинфосфату призводить до
утворення ангідриду креатину - креатиніну.
У формі креатиніну з організму людини виділяється із сечею значна частина азоту
амінокислот; у здорової людини виділення креатиніну пропорційне масі м’язових тканин і значно збільшується за умов травматичних пошкоджень м’язів.
нсфермснтативна дегідратація і дефосфорилування крсатинфосфату призводить до
утворення ангідриду креатину - креатиніну.
У формі креатиніну з організму людини виділяється із сечею значна частина азоту
амінокислот; у здорової людини виділення креатиніну пропорційне масі м’язових тканин і значно збільшується за умов травматичних пошкоджень м’язів.
 Глутатіон міститься в клітинах тваринного організму у високій концентрації (близько
5 мМ). Глутатіон оборотно перетворюється з відновленої (Г-SН) до окисленої
(Г-S-S-Г) форми, відіграючи роль буфера SН-груп.
Глутатіон міститься в клітинах тваринного організму у високій концентрації (близько
5 мМ). Глутатіон оборотно перетворюється з відновленої (Г-SН) до окисленої
(Г-S-S-Г) форми, відіграючи роль буфера SН-груп.
 Біохімічна функція глутатіону в організмі пов’язана з відновленням і детоксикацією органічних пероксидів - похідних пероксиду водню НО-ОН, у молекулі якого один (гідропероксиди) або обидва (алкілпероксиди) атоми водню заміщені на алкільні радикали:
R-O-O-H R-O-O-R
При взаємодії глутатіону з гідропероксидом утворюються нешкідливі органічні спир-
ти, що підлягають подальшому окисленню:
Реакція каталізується ферментом глутатіонпероксидазою, що містить в актив-
ному центрі атом селену (Sе).
Зворотне відновлення Г-SS-Г до Г-SH каталізується НАДФН-залежною глутатіонредуктазою.
Біохімічна функція глутатіону в організмі пов’язана з відновленням і детоксикацією органічних пероксидів - похідних пероксиду водню НО-ОН, у молекулі якого один (гідропероксиди) або обидва (алкілпероксиди) атоми водню заміщені на алкільні радикали:
R-O-O-H R-O-O-R
При взаємодії глутатіону з гідропероксидом утворюються нешкідливі органічні спир-
ти, що підлягають подальшому окисленню:
Реакція каталізується ферментом глутатіонпероксидазою, що містить в актив-
ному центрі атом селену (Sе).
Зворотне відновлення Г-SS-Г до Г-SH каталізується НАДФН-залежною глутатіонредуктазою.