КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Д в и г а т е л ь н ы й с т е р е о т и п. Понятие
|
|
|
|
Кожа
Кожа не является принадлежностью
гвигательного аппарата. Однако, реф
лекторное взаимодействие кожи и мус
кулатуры, выражающееся многообраз-
:-гы\пт феноменами (дрожание при ох-
-: -тении, повышение или понижение
«ышечного тонуса, изменение микроцир-
хуляшги) позволяет считать эти связи су
щественными. В клиническом отношении
бояее известны дерматомоторные отно-
гзекия. хотя диагностическое значение
моторно-дермальных отношений не ме-
.: - ажио. Установлено, что над участ-
вхы мышечного гипертонуса или по со-
CSZCTBV часто формируются зоны кожной
- -;г::_7гезии с одновременной пиломо-
- r-:;i реакцией. При поверхностной
пальпации, точнее при скользящем ка
сании, эти участки определяются как
препятствие скольжению пальцев —
"феномен прилипания". Механизмы
возникновения этих участков будут рас
смотрены позже.
В практическом же отношении важ
но, что "феномен прилипания" являет
ся частым диагностическим симптомом,
и что устранение кожной гипералгезии
является важным компонентом лечебных
приемов так называемой мягкой техни
ки мануальной терапии.
Кожная гипералгетическая зона (КГЗ)
характеризуется сниженной эластичнос
тью при растяжении. В этом отноше
нии можно говорить о выявлении ба
рьера (резерва движения) при исследо
вании динамического сопротивления на
растяжение.
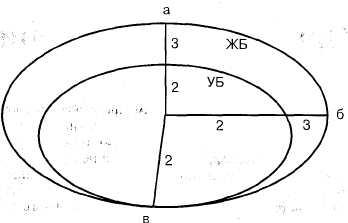
Пальпаторные ощущения при растя
жении в определенном направлении
кожи иллюстрируются на рис. 2. 7. Как
видим, активный барьер, как и при ис
следовании фасций и связок, не опре
деляется, интервал между УБ и ЖБ рас
тянут, что характеризует достаточный
резерв движения даже при КГЗ.
|
Глава 3
Организация движения
Нам предстоит познакомиться с ос
новными теоретическими сведениями
об организации и контроле движений.
В широком смысле слова под нейромо-
торной системой понимают функцио
нально-структурное единство различных
уровней нервной системы и мускула
туры. Отношения центра и периферии
(в нашем случае моторного аппарата) ор
ганизованы по типу функциональных
систем (Анохин П.К., 1975). Суть этой
организации заключается в том, что ко
мандный пункт — центр — имеет не
прерывную информацию о выполнении
задания. Это обеспечивается возвратной
(обратной) афферентацией. Следует
подчеркнуть важность афферентации в
деятельности нейромоторной системы
на всех уровнях. В деятельности центра
с помощью афферентных систем стро
ится афферентная модель будущего дви
жения со всеми его временными и про
странственными параметрами. При рас
согласовании параметров идеального,
афферентного образа движения и реаль
ного результата центр вносит соответ
ствующую поправку в эфферентную
систему. Следовательно, рассогласова
ние на входе в функциональную систе
му является условием ее активации.
Следует иметь в виду, что функциональ
ная система не является постоянной
структурой. Она может формироваться
для достижения близкой цели в малые
интервалы времени (часто бытовые яв
ления), а также для решения задач дол
госрочного, стратегического назначения,
к примеру, творчество, образование и
др. Естественно, что в этом контексте
моторика является малой частью систе
мы. Как уже говорилось, в функцио
нальной системе периферия и центр
представляют собой единство. Сейчас
нам предстоит рассмотреть основные
морфо-функциональные характеристи
ки нейромоторной системы перифери
ческой и центральной локализации.
Структурной единицей спинного моз
га является сегмент, где сгруппированы
нейроны различного назначения. Зад
ние рога и центральная часть сегмента
ориентированы на обработку сенсорных
сигналов. В нейрофизиологической ли
тературе их часто обозначают как 1-6 зоны
Рекседа. Передние рога содержат тела
эфферентных нейронов (7-9 зоны Рек
седа). Отметим, что сенсорных нейро
нов в количественном отношении на
много больше, чем эфферентных.
Мотонейроны представляют собой
обособленную группу нейронов. Наибо
лее крупные из них ос-мотонейроны с
диаметром аксона 12-22 мкм, скоростью
проведения импульса 70-120 м/с, обес
печивают выполнение произвольных
сокращений в поперечно-полосатой
мускулатуре. Потенциал действия, воз
никающий в теле нейрона в зоне аксон-
ного холмика проводится до конечного
пункта — нервно-мышечного соедине
ния (моторной бляшки). Здесь энергия
потенциала действия (ПД) расходуется
на высвобождение медиатора (в нерв
но-мышечном соединении — ацетилхо-
лин) из пресинаптического окончания,
переход его на постсинаптическую мем
брану. Затем под влиянием медиатора
происходит генерация нового ПД, но
уже на базе мышечного субстрата. Воз
никающий ПД передается на соседние
участки мышц (скорость проведения ПД
по мышце 4-6 м/с). Охватившая волна
возбуждения вызывает пространствен
ную перестройку сократительного суб
страта мышц — образование актин-мио-
зиновых комплексов, что сопровождается
Организация движения
фазическим (быстрым) сокращением
поперечно-полосатой мускулатуры.
Кроме фазического сокращения, в мус
кулатуре поддерживается постоянный
сократительный процесс, реализуемый
особыми мышечными клетками — крас
ными волокнами, в результате чего под
держивается постоянная готовность к
быстрому сокращению. Это состояние
известно как тонус мышцы. В обеспе
чении тонической активности мышц
ведущее место принадлежит малым а-мо-
тонейронам. Существует еще одна груп
па нейронов — у-эфференты, возбуж
дение которых не сопровождается ви
димым сокращением мышцы, а меня
ется состояние возбудимости проприо-
рецептора в результате деформации
участка мышцы внутри проприоцепто-
ра, т.е. интрафузального мышечного
волокна.
Следует подчеркнуть, что деятель
ность мотонейронов контролируется в
сложно организованной структуре дви
гательной системы, как в пределах сег
мента, так и за счет супраспинальных
влияний. Определенный Шеррингтоном
"принцип общего конечного пути" за
ключается в конвергенции многих ре
гулирующих влияний на мотонейроне.
Это значит, что количество афферент
ной информации должно испытывать
значительную обработку во вставочных
нейронах и на самом теле мотонейро
на, чтобы последний мог воспроизвес
ти ПД необходимой интенсивности и
точности.
Афферентный поток из мускулатуры
организован несколькими каналами.
В мышцах различают собственные про-
гтриорецепторы — нервно-мышечные
зеретена. Они соединены параллельно
основной мышце. При удлинении мыш-
_' гни возбуждаются, при ее укороче
нии активация рецептора снимается.
Установлено, что такой вид рецепции
". гнечивает центр информацией о сте-
шеяи растяжения мышцы и динамичес-
··".".грузках. Афференты этой группы
tlai заканчиваются моносинаптически на
::'··· "г.гьных клетках передних рогов,
практически не испытывая коррекции
при прохождении через сегментарный
аппарат. Это позволяет понять, что
функциональная роль такого типа про-
приорецепторов заключается в осущест
влении рефлекса на растяжение. Про-
приорецепторы мышц имеют разную
морфологию и специализацию. Извест
ны ядерно-сумчатые и ядерно-цепочные
рецепторы, а также так называемые вто
ричные окончания. Часть из них явля
ется сигнализатором статического уси
лия мышцы. Подчеркнем, что для нор
мальной функциональной активности
систем построения движения важно со
отношение проприоцепции динамичес
кого и статического характера.
Кроме мышечных рецепторов важную
роль в организации рефлекторной дея
тельности нейромоторного аппарата иг
рают сухожильные аппараты Гольджи.
Они включены последовательно (тан-
демно) с сухожилием, являются сигна
лизатором степени растягивающего уси
лия, развиваемой силы мышцы, дают
начало афферентам 1с.
Импульсация, поступающая по аффе
рентам 1а, вызывает в сегментах возбуж
дение (активацию) мотонейрона своей
мышцы и тормозит мотонейроны анта
гониста (т.е. осуществляется реципрок-
ное торможение). Афференты группы II,
начинающиеся от вторичных окончаний
веретен, путем полисинаптических вли
яний возбуждают мотонейроны сгиба
телей и тормозят мотонейроны разги
бателей.
Афференты 1о вызывают торможение
мотонейронов собственной мышцы
(собственное, т.е. аутогенное торможе
ние) и возбуждение мотонейронов ан
тагонистов.
Мышечные веретена имеют и эффе
рентную иннервацию — к ним идут ак
соны у-мотонейронов, располагающих
ся в передних рогах сегмента. В состоя
нии покоя количество импульсов из ве
ретен невелико. При растяжении мыш
цы частота импульсации из проприо-
рецепторов повышается. У первичных
окончаний частота импульсации зависит
от скорости растяжения, у вторичных —
от длины мышцы. Иначе, первичные
окончания обеспечивают динамическую
составляющую проприорецепции, вто
ричные — статическую.
Активация у-эфферентов приводит к
повышению чувствительности нервно-
мышечных веретен, т.е. рецепторов.
Установлено, что афферентация из ве
ретен может быть осуществлена не толь
ко растяжением, но и активацией у-эф
ферентов. Это означает, что рефлектор
ная возбудимость а-мотонейронов за
висит от состояния возбудимости у-мо-
тонейрона через изменение возбудимос
ти нервно-мышечного веретена. Пока
зано, что возбуждение а-мотонейронов
тоже сопровождается активацией у-эф
ферентов опять же через изменение про-
приоцепции: это так называемая а-у ко-
активация. Состояние баланса возбуди
мости рефлекторного кольца обеспечи
вается супраспинальным контролем че
рез кортикоспинальный и ретикулоспи-
нальный пути. Реализуются эти влия
ния путем изменения возбудимости как
а-, так и у-нейронов.
Таким образом, мышечные веретена
реагируют на два взаимодействия: пе
риферическое и центральное. Поэтому
в естественных условиях проприоцеп-
ция из мышц, сухожилий и суставов
испытывает сложные взаимоотношения.
Следует учесть, что проприоцепторы
суставов, связок оказывают на мотоней
роны не менее сложное влияние через
афференты различного назначения и
систему вставочных нейронов. Взаимо
действие нейронов реализуется двумя
путями — возбуждением и торможением.
Возбуждение (активация) происходит
в результате деполяризации мембраны
клетки, изменения ионной проницае
мости с последующим формированием
потенциала действия (ПД). Торможение
в своей сути тоже представляет собой ак
тивный процесс с генерацией тормозно
го потенциала, отличающегося от по
тенциала действия тем, что он локален,
т.е. не передается по нервному волокну
и имеет возможность пространственной
Мануальная медицина
и временной суммации. С точки зрения
приложения тормозных влияний на ней
рон различают пресинаптическое и
постсинаптическое торможение.
Пресинаптическое торможение осу
ществляется на входе в нейрон; в его
основе лежит деполяризация мембраны
с уменьшением величины ПД, поступа
ющего к этому нейрону из другого, в
результате чего генерация постсинапти-
ческого ПД резко угнетается.
Постсинаптическое торможение вы
зывает уменьшение величины ПД ней
рона на выходе, т.е. генерация ПД пост-
синаптической мембраной под влияни
ем тормозных потенциалов затрудняет
ся или становится невозможной. В ос
нове постсинаптического торможения
лежат механизмы гиперполяризации
мембраны синапса.
Классическим примером пресинапти-
ческого торможения является уменьше
ние сенсорного потока в заднем роге
спинного мозга, благодаря чему осу
ществляется так называемый воротный
контроль. Постсинаптическое торможе
ние, к примеру, представлено возврат
ным торможением а-мотонейрона, обес
печиваемого вставочным нейроном Рен-
шоу. В свою очередь, вставочный ней
рон активизируется деятельностью ос
новного: чем интенсивнее работа а-мо
тонейрона, тем более активен тормоз-
ный и тем большее тормозящее влия
ние испытывает мотонейрон. В сегмен
тарном аппарате известен и другой тип
торможения — двойное. В результате
торможения тормозного нейрона может
быть активизирована деятельность ос
новного.
Разумеется, основные сведения по
структурно-функциональной организа
ции сегментарного аппарата будут из
ложены без детальной характеристики
реализуемых физиологических комплек
сов, имеющих отношение к моторике.
Спинной мозг осуществляет огромное
количество рефлекторных ответов: су
хожильные рефлексы и рефлексы растя
жения имеют самое короткое латентное
время, и их физиологическое значение
Организация движения
наиболее заметно проявляется в меха
низмах фазно-тонических реакций. Бо
лее сложные рефлекторные реакции
обеспечивают координационные отно
шения, к примеру, сгибательные реф
лексы, имеющие защитное значение;
разгибательные рефлексы стопы и пр.
Еще более сложный характер имеют
ритмические рефлексы и рефлексы по
ложения (позы). В мануальной терапии
большое практическое значение имеют
шейно-тонические и вестибуло-тони-
ческие рефлексы положения Р.Магну
са. Методика их использования будет
приведена позже. Кроме рассмотренных
соматических рефлексов, выражающих
ся в активации скелетных мышц, сег
ментарный аппарат играет важную роль
в рефлекторной регуляции внутренних
органов, являясь центром висцеральных
рефлексов. Установлено, что между со
матическими и висцеральными рефлек
сами существует функциональное взаи
модействие (моторно-висцеральные
рефлексы Могендовича).
Таким образом, основная функцио
нальная единица сегментарного аппара
та — рефлекс. Как бы сложно ни была
организована рефлекторная реакция,
она строго ограничена временными и
пространственными параметрами, т.е.
жестко детерминирована местом воз
буждения, количеством активированных
нейронных групп и способом реализа
ции. Поскольку рефлекторные связи
организованы по.типу колец, включаю
щих эфферентную иннервацию аффе
рентных нейронов, этот тип организа
ции движения называют кольцевым или
коррекционным. Тем самым подчерки
вается малая пластичность и функцио
нальная жесткость описанных рефлек
торных комплексов.
Супраспинальный контроль над дея
тельностью сегментарного аппарата осу
ществляется системой нисходящих и
восходящих путей. Восходящая аффе-
рентация осуществляется системой про-
"риоцептивных путей (сознательных и
бессознательных), достигающих всех
релейных станций обработки сенсорных
сигналов. Тем самым обеспечивается
разносторонняя характеристика пара
метров движения: силы, объема, точнос
ти, плавности, целесообразности, закон
ченности, согласованности с вегетатив
ными, гуморальными, поведенческими
компонентами движения и др. Много
уровневая обработка сенсорного пото
ка составляет основу акцептора дейст
вия по П.К.Анохину, параметры кото
рого сравниваются с афферентными
параметрами реального результата дви
жения. Следует учесть, что движению
дается эмоционально-личностная, т.е.
сознательная оценка в пределах этой же
функциональной системы.
Результат обработки сенсорного по
тока является базой организации нис
ходящего контроля из вышерасположен
ных структур за рефлекторной деятель
ностью и деятельностью афферентных
систем сегментарного аппарата, обеспе
чивающего информацию с периферии.
Объектами супраспинальных влияний
являются активность мотонейронов,
вставочных нейронов, нейронов восхо
дящих систем и первичных афферент
ных волокон.
Быстропроводящие пути (кортико-,
ретикуло-, вестибулоспинальные) спо
собны возбуждать мотонейроны спин
ного мозга моносинаптически. Тем са
мым обеспечивается высокая эффектив
ность и специфичность нисходящих
команд. Установлено, что моносинап-
тические кортико-спинальные влияния
наиболее активны в мотонейронах, ин-
нервирующих дистальные мышцы ко
нечностей. Ретикуло- и вестибулоспи
нальные пути оказывают преимущест
венное влияние на мотонейроны мышц
туловища и проксимальных мышц ко
нечностей.
Нисходящие тракты оказывают влия
ние и на различные типы вставочных
нейронов спинного мозга. Наиболее
важное значение, как уже говорилось,
имеют связи с у-нейроном, результат
активации которого супраспинальной
командой имеет важное значение в ус
тановочной активности мускулатуры
в будущем движении. Реализуется же
этот тип активности изменением воз
будимости проприоцептора, предвари
тельно активированного у - н е й р о н о м.
Иначе, в афферентной системе мышцы
кодируются параметры ожидаемого дви
жения.
Нисходящие пути могут вызвать тор
можение как первичных, так и вторич
ных афферентов восходящего направле
ния. Это вызывает контроль интенсив
ности и качества сенсорного потока,
доставляемого в супраспинальные струк
туры.
Таким образом, деятельность супра-
сегментарного комплекса организации
движений интегрируется на многих
уровнях центральной нервной системы.
Основными из них являются ствол го
ловного мозга (мозжечок, вестибуляр
ный анализатор, ретикулярная форма
ция, четверохолмие), подкорковые се
рые ядра (полосатое тело, хвостатое
ядро, таламус) и сенсомоторные зоны
коры больших полушарий, включающие
передние и задние центральные изви
лины, префронтальную кору. Функци
ональные отношения между различны
ми уровнями организации движения
изучены Н.А.Бернштейном (1948).
Выделены уровни А, В, С, D, Е. Сущ
ность надсегментарной организации
движения заключается в формировании
программ движения, алгоритма двига
тельных комплексов ("мелодии движе
ни я" — по Н.А.Бернштейну). Как пра
вило, программа движения приобрета
ется большей частью индивидуальным
опытом (игра на музыкальных инстру
ментах, профессиональные двигатель
ные навыки и пр.). Конечно же, в про
грамме движения отражен и опыт ви
довой, т.е. генетический — содружест
венные движения, двигательные ком
плексы взаимодействия анализаторов,
характерные жесты, мимика и пр.
Программа движения не предусмат
ривает жесткую сцепленность этапов
выполнения, допускается изменение
параметров движения в известных пре
делах, т.е. пластичность организации
Мануальная медицина
движения. В отличие от этого, спиналь-
но-сегментарный уровень организации
движения характеризуется жесткостью
исполнения и текущего контроля за ре
ализацией каждого этапа сокращения
мышцы. Иначе, точность активации и
торможение мотонейронов сегментарно
го аппарата точно контролируется со
стоянием проприоцепции в большей
мере, чем супраспинальным контролем.
В принципе, командные супраспиналь
ные импульсы могут активировать боль
шую группу мотонейронов или даже
мотонейроны антагониста. Этим объяс
няются ошибки при заучивании новых
движений, дискоординация их при мно
гих патологических состояниях. В сег
ментарном же аппарате деятельность
отдельного мотонейрона может быть
охарактеризована возбуждением или
торможением. В наиболее общем виде
следует говорить о супрасегментарном
уровне организации как о качестве мо
торики. Сегментарный уровень обеспе
чивает ее количественную характерис
тику — силу, тонус, объем.
Постоянное взаимодействие механиз
мов программного, т.е. вероятностного,
характера организации движения с ме
ханизмами конкретных исполнителей
создает неповторимую индивидуальную
моторику как в совершенном, так и ис
каженном виде.
Клиническая практика располагает
многочисленными фактами, свидетель
ствующими о возможности быстрого
изменения функционального состояния
спинально-сегментарного аппарата,
стойкости и пластичности механизмов
супраспинального контроля. В качест
ве примера можно привести спастичес
кую кривошею.
Известно, что спустя определенный
срок от начала заболевания патологи
ческая активность центральных меха
низмов гиперкинеза головы исчезает.
Патологическая поза головы может быть
обусловлена изменениями суставно-
мышечного аппарата позвоночника,
которые лечебными приемами ману
альной терапии могут быть устранены
Организация движения
достаточно быстро. Однако, у больший-
ства больных сохраняется патологичес-
кая синкинезия — подъем плеча на сто-
роне обращенного к нему лица. Пере-
стройка этой сложившейся патологичес-
кой программы движения — патологи-
ческого двигательного стереотипа пред-
ставляет собой достаточно сложную за-
дачу.
Таким образом, в общем предметом ма-
нуальнои терапии является не только па-
тология мышц, суставов и других пери-
ферических элементов, но и патологичес-
кий двигательный стереотип. Далее мы
покажем, как под влиянием патологичес-
ких изменений на спинально-сегментар-
ном уровне организации движения мо-
жет меняться программа его построения.
Глава 4
Общая характеристика
функциональной патологии
локомоторной системы
|
|
|
|
|
Дата добавления: 2014-12-16; Просмотров: 572; Нарушение авторских прав?; Мы поможем в написании вашей работы!